Fløjlorm er segmenterede dyr med et fladt cylindrisk kropstværsnit og rækker af ustrukturerede kropsbøjninger kendt som oncopods eller lobopods (uformelt: stubfødder). Dyrene vokser til mellem 0,5 og 20 cm (.2 til 8 tommer), hvor gennemsnittet er omkring 5 cm (2 tommer) og har mellem 13 og 43 par ben. Deres hud består af talrige, fine tværgående ringe og er ofte iøjnefaldende farvet orange, rød eller brun, men nogle gange også lysegrøn, blå, guld eller hvid og lejlighedsvis mønstret med andre farver. Segmentering er udadtil ubemærket og kan identificeres ved den regelmæssige afstand mellem par af ben og i det regelmæssige arrangement af hudporer, udskillelsesorganer og koncentrationer af nerveceller. De enkelte kropssektioner er stort set uspecialiserede; selv hovedet udvikler sig kun lidt anderledes end mavesegmenterne. Segmentering er tilsyneladende specificeret af det samme gen som i andre grupper af dyr og aktiveres i hvert tilfælde under embryonal udvikling ved den bageste kant af hvert segment og i vækstområdet for stubben. Selvom onychophorans falder inden for protostomegruppen, har deres tidlige udvikling en deuterostomebane (med munden og anus dannet separat); denne bane er skjult af de ret sofistikerede processer, der forekommer i den tidlige udvikling.
AppendagesEdit
stubfødderne, der kendetegner fløjlormene, er koniske, baggy vedhæng i kroppen, som er internt hule og ikke har led. Selvom antallet af fødder kan variere betydeligt mellem arter, er deres struktur stort set meget ens. Stivhed tilvejebringes af det hydrostatiske tryk af deres væskeindhold, og bevægelse opnås normalt passivt ved strækning og sammentrækning af dyrets hele krop. Imidlertid kan hvert ben også forkortes og bøjes af indre muskler. På grund af manglen på ledd kan denne bøjning finde sted på ethvert tidspunkt langs benets sider. I nogle arter findes to forskellige organer inden for fødderne:
- Crural kirtler er placeret ved benets skulder og strækker sig ind i kropshulen. De åbner udad på crural papiller-små vorte-lignende bump på maven side af benet—og udskiller kemiske messenger materialer kaldet feromoner. Deres navn kommer fra den latinske cruralis, der betyder “af benene”. vesikler er poser placeret på maven side af benet, som kan vendes og sandsynligvis tjene i vandabsorption. De tilhører familien Peripatidae og er navngivet fra koks, det latinske ord for “hofte”.

på hver fod er et par udtrækkelige, hærdede (sclerotiserede) chitinklør, som giver dens videnskabelige navn: onychophora er afledt af de græske onyches, “kløer”; og pherein, “at bære”. Ved bunden af klørne er der tre til seks spiny “puder”, hvorpå benet sidder i sin hvileposition, og hvorpå dyret går over glatte underlag. Klørne bruges hovedsageligt til at få et fast fodfæste på ujævnt terræn. Hver klo består af tre stablede elementer, som russiske nesting dukker. Den yderste er skuret under ecdysis, som udsætter det næste element i — som er fuldt dannet, så behøver ikke tid til at hærde, før det bruges. (Denne karakteristiske konstruktion identificerer mange tidlige kambriske fossiler som tidlige udløbere af onychophoran-slægten.) Bortset fra benparene er der yderligere tre kropsbøjlersom er i hovedet og omfatter tre segmenter:
- på det første hovedsegment er et par slanke antenner, der tjener i sensorisk opfattelse. De svarer sandsynligvis ikke direkte til leddyrets antenner, men måske snarere med deres “læber” eller labrum. Ved deres base findes et par enkle øjne, undtagen i nogle få blinde arter. Foran disse er der i mange Australske arter forskellige fordybninger, hvis funktion endnu ikke er klar. Det ser ud til, at i det mindste nogle arter tjener disse til overførsel af sædcellepakker (spermatoforer).
- på mavesiden af det andet hovedsegment er labrum, en mundåbning omgivet af følsomme “læber”. I fløjlormene er denne struktur en muskuløs udvækst i halsen, så på trods af sit navn er det sandsynligvis ikke homologt med Arthropodas labrum og bruges til fodring. Dybt inde i mundhulen ligger de skarpe, halvmåneformede” kæber ” eller mandibler, som er stærkt hærdede og ligner føddernes kløer, som de er serielt homologe med; tidligt i udviklingen har kæbebøjlerne en position og form svarende til de efterfølgende ben. Kæberne er opdelt i indre og ydre mandibler, og deres konkave overflade bærer fine dentikler. De bevæger sig bagud og fremad i længderetningen, rive byttet fra hinanden, tilsyneladende bevæget i den ene retning af muskulatur og den anden ved hydrostatisk tryk. Klørne er lavet af sklerotiseret Kurt-chitin, forstærket med phenoler og kinoner og har en ensartet sammensætning – bortset fra at der er en højere koncentration af calcium mod spidsen, hvilket formodentlig giver større styrke.
overfladen af mandibles er glat, uden ornamentik. Neglebåndet i mandiblerne (og klørne) adskiller sig fra resten af kroppen. Det har en indre og ydre komponent; den ydre komponent har kun to lag (hvorimod krops neglebånd har fire), og disse ydre lag (især den indre epicuticle) er dehydreret og stærkt garvet, hvilket giver sejhed.
Slime kirtlerrediger
på det tredje hovedsegment til venstre og højre for munden er to åbninger betegnet “orale papiller”. Inden for disse er et par store, stærkt internt forgrenede slimkirtler. Disse ligger omtrent i midten af kroppen og udskiller en slags mælkehvid slim, som bruges til at fange bytte og til defensive formål. Nogle gange udvides den forbindende “slimleder” til et reservoir, som kan buffer præproduceret slim. Selve slimkirtlerne er sandsynligvis modificerede crural kirtler. Alle tre strukturer svarer til en evolutionær oprindelse i benparene i de andre segmenter.
hud og musklerrediger
i modsætning til leddyrene har fløjlorm ikke et stift eksoskelet. I stedet fungerer deres væskefyldte kropshulrum som et hydrostatisk skelet på samme måde som mange ikke-relaterede bløddyr, der er cylindrisk formede, for eksempel havanemoner og forskellige orme. Tryk på deres ukomprimerbare indre kropsvæske på kropsvæggen giver stivhed, og musklerne er i stand til at handle imod det. Kropsvæggen består af en ikke-cellulær ydre hud, cuticula; et enkelt lag af epidermisceller, der danner en indre hud; og under dette, normalt tre lag muskler, som er indlejret i bindevæv. Cuticula er omkring en mikrometer tyk og dækket med fine villi. I sammensætning og struktur ligner den cuticula af leddyrene, der består af Kurt-chitin og forskellige proteiner, selvom de ikke indeholder kollagen. Det kan opdeles i en ekstern epicuticula og en intern procuticula, som selv består af ekso – og endo-cuticula. Denne struktur på flere niveauer er ansvarlig for den høje fleksibilitet i den ydre hud, som gør det muligt for fløjlormen at presse sig ind i de smaleste sprækker. Selvom udadtil vandafvisende, er cuticula ikke i stand til at forhindre vandtab ved åndedræt, og som følge heraf kan fløjlormme kun leve i mikroklimaer med høj luftfugtighed for at undgå udtørring. Overfladen på neglebåndet er spredt med adskillige fine papiller, hvoraf den største bærer synlige villilignende følsomme børster. Papillerne selv er dækket af små skalaer, der giver huden et fløjlsagtigt udseende (hvorfra det almindelige navn sandsynligvis er afledt). Det føles også som tørt fløjl at røre ved, som dets vandafvisende natur er ansvarlig for. Fældning af huden (ecdysis) finder sted regelmæssigt omkring hver 14.dag induceret af hormonet ecdyson. Den indre overflade af huden bærer et sekskantet mønster. Ved hver moult erstattes skurhuden af epidermis, som ligger umiddelbart under den; i modsætning til cuticula består dette af levende celler. Under dette ligger et tykt lag af bindevæv, der primært består af kollagenfibre, der er justeret enten parallelt eller vinkelret på kroppens længdeakse. Farvningen af Onychophora genereres af en række pigmenter. Opløseligheden af disse pigmenter er en nyttig diagnostisk karakter: i alle leddyr og tardigrader er kropspigmentet opløseligt i ethanol. Dette gælder også for Peripatidae, men i tilfælde af Peripatopsidae er kropspigmentet uopløseligt i ethanol.
i bindevævet ligger tre kontinuerlige lag af uspecialiseret glat muskelvæv. Det relativt tykke ydre lag er sammensat af ringformede muskler og det tilsvarende voluminøse indre lag af langsgående muskler. Mellem dem ligger tynde diagonale muskler, der vind bagud og fremad langs kroppens akse i en spiral. Mellem de ringformede og diagonale muskler findes fine blodkar, der ligger under hudens overfladisk genkendelige tværgående ringe og er ansvarlige for de pseudosegmenterede markeringer. Under det indre muskellag ligger kropshulrummet. I tværsnit er dette opdelt i tre regioner med såkaldte dorso-ventrale muskler, der løber fra midten af underbukken til kanterne på oversiden: en central midsektion og til venstre og højre, to sideområder, der også inkluderer benene.
Cirkulationredit
kropshulrummet er kendt som et “pseudocoel” eller hæmocoel. I modsætning til en ægte coelom er en pseudocoel ikke helt lukket af et cellelag afledt af den embryonale mesoderm. En coelom er imidlertid dannet omkring gonaderne og affald-eliminere nefridier. Som navnet haemocoel antyder, er kropshulrummet fyldt med en blodlignende væske, hvor alle organerne er indlejret; på denne måde kan de let forsynes med næringsstoffer, der cirkulerer i blodet. Denne væske er farveløs, da den ikke indeholder pigmenter; af denne grund tjener det kun en begrænset rolle i ilttransport. To forskellige typer blodceller (eller hæmocytter) cirkulerer i væsken: amoebocytter og nefrocytter. Amoebocytterne fungerer sandsynligvis i beskyttelse mod bakterier og andre fremmedlegemer; i nogle arter spiller de også en rolle i reproduktion. Nephrocytter absorberer toksiner eller omdanner dem til en form, der er egnet til eliminering af nefridia. Haemocoel er opdelt af en vandret skillevæg, membranen, i to dele: perikardial sinus langs ryggen og perivisceral sinus langs maven. Førstnævnte omslutter det rørlignende hjerte, og sidstnævnte, de andre organer. Membranen er perforeret mange steder, hvilket muliggør udveksling af væsker mellem de to hulrum. Hjertet selv er et rør af ringformede muskler bestående af epitelvæv, med to laterale åbninger (ostia) pr. Selvom det ikke vides, om bagenden er åben eller lukket, åbner den forfra direkte ind i kropshulen. Da der ikke er nogen blodkar, bortset fra de fine kar, der løber mellem muskelagene i kropsvæggen og et par arterier, der forsyner antennerne, kaldes dette en åben cirkulation. Tidspunktet for pumpeproceduren kan opdeles i to dele: diastol og systole. Under diastolen strømmer blod gennem Ostia fra perikardial sinus (hulrummet indeholdende hjertet) ind i hjertet. Når systolen begynder, lukker ostia og hjertemusklerne trækker sig indad, hvilket reducerer hjertets volumen. Dette pumper blodet fra den forreste ende af hjertet ind i perivisceral sinus, der indeholder organerne. På denne måde forsynes de forskellige organer med næringsstoffer, før blodet endelig vender tilbage til perikardial sinus via perforeringerne i membranen. Ud over Hjertets pumpevirkning har kropsbevægelser også indflydelse på cirkulationen.
RespirationEdit
iltoptagelse sker i en grad via simpel diffusion gennem hele kropsoverfladen, hvor de koksale vesikler på benene muligvis er involveret i nogle arter. Af største betydning er imidlertid gasudveksling via fine forgrenede rør, luftrøret, der trækker ilt fra overfladen dybt ind i de forskellige organer, især hjertet. Væggene i disse strukturer, som er mindre end tre mikrometer tykke i deres helhed, består kun af en ekstremt tynd membran, gennem hvilken ilt let kan diffundere. Luftrøret stammer fra små åbninger, spiraklerne, som selv er samlet i dentlignende udsparinger af den ydre hud, atrierne. Antallet af således dannede” luftrørsbundter ” er i gennemsnit omkring 75 pr. kropssegment; de akkumuleres tættest på bagsiden af organismen. I modsætning til leddyrene er fløjlormene ikke i stand til at kontrollere åbningerne i deres luftrør; luftrøret er altid åbent, hvilket medfører betydeligt vandtab under tørre forhold. Vand går tabt dobbelt så hurtigt som i regnorme og fyrre gange hurtigere end i larver. Af denne grund er fløjlorme afhængige af levesteder med høj luftfugtighed.
Fordøjelsedit
fordøjelseskanalen begynder lidt bag hovedet, munden ligger på undersiden lidt fra kroppens forreste punkt. Her kan bytte mekanisk adskilles af mandiblerne med deres dækning af fine tandhjul. To spytkirtler udledes via en fælles leder i den efterfølgende” hals”, som udgør den første del af fortarmen. Spyt, som de producerer, indeholder slim og hydrolytiske stoffer, som initierer fordøjelsen i og uden for munden. Halsen i sig selv er meget muskuløs og tjener til at absorbere den delvist flydende mad og til at pumpe den via spiserøret, der danner den bageste del af fortarmen, ind i tyndtarmen. I modsætning til den forreste tarm er dette ikke foret med en neglebånd, men består i stedet kun af et enkelt lag epitelvæv, som ikke udviser iøjnefaldende indrykning, som det findes hos andre dyr. Når de kommer ind i tyndtarmen, overtrækkes fødepartikler med en slimbaseret peritrofisk membran, der tjener til at beskytte tarmens foring mod skader fra skarpe kanter. Tarmepitelet udskiller yderligere fordøjelsessymer og absorberer de frigivne næringsstoffer, selvom størstedelen af fordøjelsen allerede har fundet sted eksternt eller i munden. Ufordøjelige rester ankommer i den bageste tarm eller rektum, som igen er foret med en neglebånd, og som åbner ved anus, placeret på undersiden nær bagenden.
i næsten hvert segment er et par udskillelsesorganer kaldet nefridia, som er afledt af coelomvæv. Hver består af en lille pose, der er forbundet via en flagelleret leder kaldet en nephridioduct, til en åbning i bunden af det nærmeste ben kendt som en nephridiopore. Posen er optaget af specielle celler kaldet podocytter, som letter ultrafiltrering af blodet gennem skillevæggen mellem hæmokoelom og nefridium. Sammensætningen af urinopløsningen modificeres i nephridioduct ved selektiv genvinding af næringsstoffer og vand og ved isolering af gift og affaldsmaterialer, før den udskilles til omverdenen via nephridiopore. Det vigtigste nitrogenholdige udskillelsesprodukt er den vanduopløselige urinsyre; dette kan udskilles i fast tilstand med meget lidt vand. Denne såkaldte uricoteliske udskillelsestilstand repræsenterer en tilpasning til livet på land og den tilhørende nødvendighed af at håndtere økonomisk med vand. Et par tidligere nefridier i hovedet blev sekundært omdannet til spytkirtlerne, mens et andet par i det sidste segment af mandlige prøver nu tjener som kirtler, der tilsyneladende spiller en rolle i reproduktion.
SensationEdit
hele kroppen, inklusive stubben, er fyldt med adskillige papiller: vorte fremspring reagerer på berøring, der bærer en mekanoreceptiv børste ved spidsen, som hver også er forbundet med yderligere sensoriske nerveceller, der ligger under. Mundpapillerne, udgangene til slimkirtlerne, har sandsynligvis også en vis funktion i sensorisk opfattelse. Sensoriske celler kendt som” sensills “på” læber ” eller labrum reagerer på kemiske stimuli og er kendt som kemoreceptorer. Disse findes også på de to antenner, som synes at være fløjlormens vigtigste sanseorganer.
undtagen i nogle få (typisk underjordiske) arter ligger et enkelt konstrueret øje (ocellus) bag hver antenne, lateralt, lige under hovedet. Dette består af en chitinøs kuglelinse, en hornhinde og en nethinde og er forbundet til midten af hjernen via en optisk nerve. Nethinden består af adskillige pigmentceller og fotoreceptorer; sidstnævnte er let modificerede flagellerede celler, hvis flagellummembraner bærer et lysfølsomt pigment på deres overflade. De rhabdomeriske øjne på Onychophora menes at være homologe med median ocelli af leddyr; dette antyder, at den sidste fælles forfader til leddyr muligvis kun har haft median ocelli.Innerveringen viser imidlertid, at homologien er begrænset: onychophoras øjne dannes bag antennen, mens det modsatte er tilfældet i leddyr.
Reproduktionredit
begge køn har par gonader, der åbner via en kanal kaldet en gonoduct i en fælles kønsåbning, gonoporen, som er placeret på den bageste ventrale side. Både gonaderne og gonoduct er afledt af ægte coelomvæv.
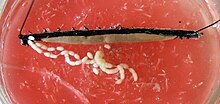
hos kvinder er de to æggestokke forbundet i midten og til den vandrette membran. Gonodukten fremstår forskelligt afhængigt af om arten er levende bærende eller æglægning. I levende bærende arter opdeles hver udgangskanal i en slank ovidukt og en rummelig “livmoder”, livmoderen, hvor embryonerne udvikler sig. Den enkelte vagina, som begge uteri er forbundet med, løber udad til gonoporen. Hos æglægningsarter, hvis gonoduct er ensartet konstrueret, ligger kønsåbningen ved spidsen af et langt æglægningsapparat, ovipositoren. Hunnerne af mange arter har også et sædopbevaringssted kaldet beholderen seminis, hvor sædceller fra mænd kan opbevares midlertidigt eller i længere perioder.
hanner har to separate testikler sammen med den tilsvarende sædblære (vesicula seminalis) og udgangskanal (vasa efferentia). De to vasa efferentia forenes til en fælles sædkanal, vas deferens, som igen udvides gennem ejakulatorisk kanal for at åbne ved gonopore. Direkte ved siden af eller bag dette ligger to par specielle kirtler, som sandsynligvis tjener en eller anden hjælpeforplantningsfunktion; de bageste kirtler er også kendt som analkirtler. En penislignende struktur er hidtil kun fundet hos mænd af slægten Paraperipatus, men er endnu ikke blevet observeret i aktion.
der er forskellige parringsprocedurer: I nogle arter deponerer mænd deres spermatofor direkte i kvindens kønsorganåbning, mens andre deponerer det på kvindens krop, hvor neglebåndet vil kollapse og lade sædcellerne migrere ind i kvinden. Der er også Australske arter, hvor hanen placerer deres spermatofor oven på hovedet, som derefter presses mod kvindens kønsorganer. I disse arter har hovedet udførlige strukturer som pigge, rygsøjler, hule styletter, pits og depressioner, hvis formål er enten at holde sædcellerne og / eller hjælpe med sædoverførslen til kvinden. Hannerne af de fleste arter udskiller også et feromon fra kirtler på undersiden af benene for at tiltrække hunner.