kaksi jännittävintä ja monipuolisinta viimeisen 30 vuoden aikana suunniteltua geenivälinettä ovat Cre-lox-ja FLP-FRT-teknologiat. Molempien avulla geenien ilmentymisen sijaintia ja ajoitusta voidaan säädellä tarkasti. Tässä artikkelissa esitellään lyhyesti, miten nämä kaksi järjestelmää toimivat ja niitä voidaan käyttää hiirimalleissa.
Cre-lox-järjestelmä
Cre-lox-tekniikka otettiin käyttöön 1980-luvulla (Sauer ja Henderson 1988; Sternberg ja Hamilton 1981) ja sen patentoi DuPont Pharmaceuticals. Sitä on käytetty menestyksekkäästi hiivoissa, kasveissa, nisäkkäiden soluviljelmissä ja hiirissä (Araki et al. 1987). Se perustuu P1-bakteriofagisyklization rekombination (Cre) – rekombinaasigeenin (cre) kykyyn saada aikaan rekombinaatio loxP-alueiden parien välillä. Tällainen rekombinaatio ”Cre-lox” – hiirellä (KS. alla) voi joko aktivoida tai inaktivoida kiinnostavan geenin.
käyttääkseen Cre-lox-tekniikkaa tutkijan on tuotettava Cre-lox-hiiri, tyypillisesti jalostamalla Cre-hiiri loxP-hiireksi. Cre-hiiri sisältää Cre-rekombinaasi-transgeenin kudoskohtaisen promoottorin ohjauksessa; loxP-hiiri sisältää kaksi loxP-aluetta, jotka reunustavat kiinnostavan genomijalan, ”floxedin” lokuksen. Tyypillisesti Cre-ja loxP-hiiriä valmistetaan siirtogeenisellä tekniikalla (Nagy 2000). Cre-hiiret voidaan promoottoreista ja muista niiden rakentamiseen käytetyistä kontrolleista riippuen suunnitella ilmaisemaan Cre-rekombinaasia vain tietyin edellytyksin, mukaan lukien seuraavat: tietyissä kudoksissa, kun hiiren ruokavaliota täydennetään aineilla, kuten doksisykliini, tetrasykliini, RU486 ja tamoksifeeni (Brocard et al. 1998; Kellendonk ym. 1999; Utomo ym. 1999) ja tietyissä kehitysvaiheissa.
Cre-lox-hiiren loxP-alueiden sijainnista ja suunnasta riippuen Cre-rekombinaasi voi aloittaa floksoidun lokuksen poistoja, inversioita ja translokaatioita (Nagy 2000) (Kuva. 1).
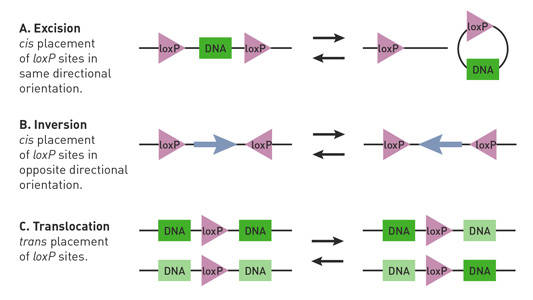 Fig. 1.
Fig. 1.
Cre-lox-reaktioihin vaikuttaa loxP-paikkojen suunta ja sijainti. Pareittain loxP sivustoja (kolmiot) on suuntaisuus ja voidaan sijoittaa cis (sama DNA-juoste) tai trans (eri DNA-juosteet) järjestely. (A) jos loxP-alueet sivuavat DNA-segmenttiä (suorakulmiota) cis-järjestelyssä ja ovat suuntautuneet samaan suuntaan, Cre-rekombinaasi välittää segmentin poistoa tai kiertokulkua. (B) Jos loxP-alueet sivuavat DNA-segmenttiä cis-järjestelyssä ja ovat suuntautuneet vastakkaisiin suuntiin, Cre-rekombinaasi välittää janan inversiota. C) Jos loxP-alueet sijaitsevat eri DNA-säikeillä ja ovat suuntautuneet samaan suuntaan, Cre-rekombinaasi välittää segmentin translokaation.
yleensä Cre-ja loxP-kannat kehittyvät itsenäisesti ja sitten risteytyvät. Monia eri Cre-kantoja, joista jokainen sisältää Cre-transgeenin eri kudosspesifisen promoottorin johdolla, voidaan risteytyä yhden loxP-kannan kanssa. Riippuen siitä, mitkä kannat pariutuvat, voidaan rakentaa erilaisia Kre-välitteisiä mallijärjestelmiä, mukaan lukien transgeeniset, tyrmäykset, hypomorfit, korjattavissa olevat hypomorfit, kromosomipoikkeavuudet ja ruokavalion aiheuttamat mutantit. Cre-ja loxP-kantoja sekoittamalla ja sovittamalla tutkija voi tutkia geenin vaikutuksia kudosspesifisillä ja kehitysvaihespesifisillä tavoilla, jotka olivat aiemmin mahdottomia.
FLP-FRT-järjestelmä
FLP-FRT-järjestelmä muistuttaa Cre-lox-järjestelmää ja on yleistymässä hiiripohjaisessa tutkimuksessa. Siinä käytetään Saccharomyces cerevisiae-hiivasta (Sadowski 1995) saatua flippase (FLP) – rekombinaasia. FLP tunnistaa FLP-rekombinaasi-kohdesekvenssiparin (FRT), joka kiertää kiinnostavan genomialueen.
Araki K, Imaizumi T, Okuyama K, Oike Y, Yamamura K. 1997. Rekombinaation tehokkuus Cre-ohimenevän ilmentymisen avulla alkion kantasoluissa: eri promoottoreiden vertailu. J Biochem (Tokio) 122: 977-82.
Brocard J, Feil R, Chambon P, Metzger D. 1998 kimeerinen Cre-rekombinaasi, joka indusoituu synteettisesti, mutta ei glukokortikoidireseptorin luonnollisten ligandien vaikutuksesta. Nukleiinihapot Res 26: 4086-90.
JAX toteaa. 1999. Nih, Jackson Laboratory ja DuPont Pharmaceuticals allekirjoittavat Cre-lox technology use-sopimukset. JAX NOTES 476: 4.
Kellendonk C, Tronche F, Reichardt HM, Schutz G. 1999. Glukokortikoidireseptorin mutageneesi hiirillä. J Steroidi Biochem Mol Biol 69: 253-9.
Nagy A. 2000. Cre-rekombinaasi: genomin räätälöinnin universaali reagenssi. Mooseksen kirja 26: 99-109.
Sadowski P. 1995. Saccharomyces cerevisiae-organismin 2-µm plasmidin FLP-rekombinaasi. Prog Nucleic Acid Res Mol Biol 51: 53-91.
Sauer B, Henderson N. 1988. Paikkaspesifinen DNA-rekombinaatio nisäkässoluissa bakteriofagi P1: n Cre-rekombinaasilla. Proc Natl Acad Sci U S A 85: 5166-70.
Sternberg N, Hamilton D. 1981. Bakteriofagi P1-paikkasidonnainen rekombinaatio. I rekombinaatio loxP-alueiden välillä. J Mol Biol 150: 467-86.
Utomo AR, Nikitin AY, Lee WH. 1999. Aika -, tila-ja solutyyppispesifinen kre-välitteisen DNA-rekombinaation kontrolli siirtogeenisillä hiirillä. Nat Biotechnol 17: 1091-6.