最後の30年に設計されている最も刺激的で、最も多目的な遺伝用具の二つはCre-loxおよびFLP-FRTの技術である。 両方とも、遺伝子発現の位置およびタイミングを密接に調節することを可能にする。 この記事では、2つのシステムがどのように機能し、マウスモデルで使用できるかを簡単に説明します。Cre-loxシステム
Cre-lox技術は1980年代に導入され(Sauer and Henderson1988;Sternberg and Hamilton1981)、デュポン製薬によって特許を取得しました。 これは、酵母、植物、哺乳動物細胞培養物およびマウスにおいて成功裏に適用されている(Araki et al. 1987). これは、p1バクテリオファージ環化組換え(Cre)リコンビナーゼ遺伝子(cre)がloxP部位の対間の組換えをもたらす能力に基づいている。 「Cre−lox」マウス(下記参照)におけるそのような組換えは、目的の遺伝子を活性化または不活性化することができる。Cre-lox技術を使用するには、研究者は、通常、creマウスをloxPマウスに繁殖させることによって、Cre-loxマウスを生産する必要があります。
cre-lox技術を使用するには、cre-loxマ Creマウスは、組織特異的プロモーターの指示の下でCreリコンビナーゼ導入遺伝子を含み、loxPマウスは、関心のゲノムセグメント、”フロックス”遺伝子座に隣接する二つのloxPサイトを含んでいます。 典型的には、CreおよびloxPマウスは、遺伝子導入技術(Nagy2 0 0 0)を使用することによって産生される。 プロモーターおよびそれらを構築するために使用される他の調節制御に応じて、Creマウスは、以下を含む特定の条件下でのみCre組換え酵素を発現するよ: 特定の組織では、マウスの食事にドキシサイクリン、テトラサイクリン、RU4 8 6およびタモキシフェンなどの物質が補充される場合(Brocard e t a l. 1 9 9 8;Kellendonk e t a l. 1 9 9 9;Utomo e t a l. 1999年)と特定の発達段階の間に。 Cre-loxマウスにおけるloxP部位の位置および向きに依存して、Cre組換え酵素は、floxed遺伝子座の欠失、反転、および転座を開始することができる(Nagy2000)(図10)。
Cre-loxマウ 1).
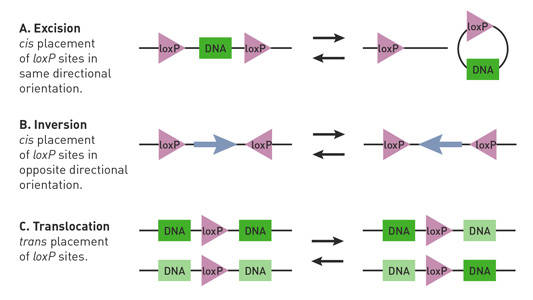 図。 1.
図。 1.
Cre-lox反応は、loxP部位の配向および位置によって影響される。 対になったloxP部位(三角形)は方向性を有し、cis(同じDNA鎖)またはtrans(異なるDNA鎖)配置に配置され得る。 (A)loxP部位がcis配列でDNAセグメント(長方形)に隣接し、同じ方向に配向している場合、Cre組換え酵素はセグメントの切除または環状化を仲介する。 (B)loxP部位がcis配列でDNAセグメントに隣接し、反対方向に配向している場合、Creリコンビナーゼはセグメントの反転を仲介する。 (C)loxP部位がDNAの異なる鎖上に位置し、同じ方向に配向している場合、Cre組換え酵素は、セグメントの転座を媒介する。
通常、Cre株とloxP株は独立して開発され、その後交配されます。
それぞれが異なる組織特異的プロモーターの指示の下でCre導入遺伝子を含有する多くの異なるCre株を、単一のloxP株と交配させることができる。 どの株が交配されているかに応じて、トランスジェニック、ノックアウト、hypomorphs、修復可能なhypomorphs、染色体異常および食事誘発変異体を含む様々なCreを介したモデ 実際には、Cre株とloxP株を混合して照合することにより、研究者は、以前は不可能であった組織特異的および発達段階特異的な方法で遺伝子の効果を調FLP-FRTシステムはCre-loxシステムに似ており、マウスベースの研究でより頻繁に使用されるようになってきています。
FLP-FRTシステムは、Cre-loxシステムに似ています。
flp-FRTシステムは、Cre-loxシステムに類似しています。 これは、酵母Saccharomyces cerevisiae(Sadowski1995)に由来するflippase(FLP)リコンビナーゼを使用することを含む。 FLPは、関心のあるゲノム領域に隣接するFLPリコンビナーゼターゲット(FRT)配列のペアを認識します。
荒木K,今泉T,奥山K,大池Y,山村K.1997. 胚性幹細胞におけるCre一過性発現による組換え効率:種々のプロモーターの比較。 Jバイオケム(東京)122:977-82。
Brocard J,Feil R,Chambon P,Metzger D.1998a chimeric Cre recombinase inducible by synthetic,but not by natural ligands of glucocorticoid receptor. Nucleic Acids Res2 6:4 0 8 6−9 0.
JAXノート。 1999. NIH、Jackson Laboratory、およびDuPont Pharmaceuticalsは、Cre-lox技術使用契約に署名しています。 JAXノート476:4。
Kellendonk C,Tronche F,Reichardt HM,Schutz G.1999. マウスにおけるグルココルチコイド受容体の変異誘発。 JステロイドBiochem Mol Biol69:253-9.
Nagy A.2000. Creリコンビナーゼ:ゲノム仕立てのための普遍的な試薬。 創世記第26章99-109節。
サドフスキP.1995。 Saccharomyces cerevisiaeの2-μ mプラスミドのFlpリコンビナーゼ。 Prog Nucleic Acid Res Mol Biol5 1:5 3−9 1.
Sauer B,Henderson N.1988. バクテリオファージP1のCreリコンビナーゼによる哺乳類細胞における部位特異的DNA組換え。 Proc Natl Acad Sci U S A85:5166-70.
Sternberg N,Hamilton D.1981. バクテリオファージP1部位特異的組換え。 I.loxPサイト間の組換え。 J Mol Biol1 5 0:4 6 7−8 6.
Utomo AR,Nikitin AY,Lee WH. 1999. トランスジェニックマウスにおけるCre媒介DNA組換えの時間的、空間的、および細胞型特異的制御。 ナターシャ17:1091-6.