ベルベットのワームは、平らな円筒形の体の断面とoncopodsまたはlobopods(非公式:スタブ足)として知られている非構造化された体の付属物の行を持つセグメ 動物は0.5と20センチメートル(の間に成長します。であり、平均は約5cm(2インチ)であり、1 3対〜4 3対の脚を有する。 彼らの皮膚は多数の、細かい横方向のリングで構成されており、しばしば目立たないようにオレンジ、赤または茶色に着色されていますが、時には明るい緑、青、金または白、時には他の色でパターン化されています。 セグメンテーションは外側に目立たず、足のペアの規則的な間隔と皮膚孔、排泄器官および神経細胞の濃度の規則的な配置によって識別可能である。 個々のボディセクションは主としてunspecialised;頭部は腹部の区分と少しだけ別様に成長する。 セグメンテーションは、明らかに動物の他のグループと同じ遺伝子によって指定され、それぞれの場合、胚発生中、各セグメントの後縁およびスタブ足の成長ゾーンで活性化される。 Onychophoransはプロトストームグループ内に分類されますが、その初期の発達は(口と肛門が別々に形成されている)重水素軌道を持っています。
AppendagesEdit
ベルベットのワームを特徴付けるスタブ足は、内部的に中空であり、関節を持たない体の円錐形のだぶだぶの付属物です。 足の数は種によってかなり異なることがありますが、その構造は基本的に非常に似ています。 剛性は、それらの流体内容物の静水圧によって提供され、運動は通常、動物の全身の伸張および収縮によって受動的に得られる。 しかし、各脚はまた、内部の筋肉によって短縮され、曲げることができる。 関節の欠如のために、この曲げは脚の両側に沿った任意の時点で起こることができる。 いくつかの種では、足の中に二つの異なる器官が見られます:
- Crural腺は脚の肩に位置し、体腔に伸びています。 それらはcrural乳頭—足の腹側面の小さい疣そっくりの隆起-で外側に開き、フェロモンと呼ばれる化学メッセンジャー材料を分泌する。 彼らの名前は、”足の”を意味するラテン語のcruralisから来ています。
- Coxal小胞は、脚の腹側に位置するパウチであり、これはevertedされ、おそらく吸水に役立つ。 彼らは家族Peripatidaeに属し、coxa、”ヒップ”のためのラテン語から命名されています。

各足に格納式、硬化(硬化)キチン爪のペアであり、分類群にその学名を与える:Onychophoraギリシャ語のonyches、”爪”から派生しています。 爪の基部には、脚がその休息位置に座っており、動物が滑らかな基材の上を歩く三から六の棘のある”クッション”があります。 爪は主に不均一な地形でしっかりとした足場を得るために使用されます。 それぞれの爪は、ロシアの入れ子人形のような3つの積み重ねられた要素で構成されています。 最も外側は脱脂の間に流され、完全に形成された次の要素を露出させるので、使用する前に硬化する時間を必要としない。 (この独特の構造は、多くの初期のカンブリア紀の化石をオニコホランの系統の初期の分派として同定している。)足のペアから離れて、頭にあり、三つのセグメントを含む三つのさらなる身体付属物があります:
- 最初の頭のセグメントには、感覚知覚に役立つ細い触 彼らはおそらく節足動物の触角に直接対応するのではなく、おそらくむしろ彼らの”唇”またはラブラムに対応しています。 彼らの基部には、いくつかの盲目の種を除いて、単純な目のペアが見つかりました。 これらの前には、多くのオーストラリアの種では、さまざまなディンプルがあり、その機能はまだ明らかではありません。 少なくともいくつかの種では、これらは精子細胞パッケージ(spermatophores)の移動に役立つようである。
- 第二の頭のセグメントの腹側にlabrum、敏感な”唇”に囲まれた口の開口部です。 ベルベットのワームでは、この構造は喉の筋肉の成長であるため、その名前にもかかわらず、おそらく節足動物の唇と相同ではなく、摂食に使用されます。 口腔内の奥深くには、鋭い三日月形の”顎”、または下顎があり、それらは強く硬化し、足の爪に似ており、それらは連続的に相同であり、発達の初期には、顎の付属物は、その後の脚に類似した位置および形状を有する。 顎は内部および外的な下顎に分けられ、凹面の表面は良い歯質に耐えます。 彼らは長手方向に前後に移動し、獲物を引き裂き、明らかに筋肉によって一方向に移動し、静水圧によって他の方向に移動する。 爪は硬化したα-キチンでできており、フェノールとキノンで補強されており、均一な組成を持っていますが、先端に向かってカルシウムの濃度が高く、おそらくより大きな強度を与えていることを除いています。
下顎骨の表面は滑らかで、装飾はありません。 下顎(および爪)のキューティクルは、体の残りの部分とは異なります。 それは内側と外側のコンポーネントを持っています; 外の部品にちょうど2つの層が(ボディクチクラに4がある一方)あり、これらの外の層(特に内部のepicuticle)は脱水され、強く日焼けし、靭性を与えます。
スライム腺編集
口の左右にある第三の頭部セグメントには、”口腔乳頭”と指定された二つの開口部があります。 これらの中には、大きな、重く内部的に分岐したスライム腺のペアがあります。 これらは体のほぼ中央にあり、一種の乳白色のスライムを分泌し、獲物を捕らえるために、そして防御目的のために使用されます。 時には、接続する”スライム導体”は、予め生成されたスライムを緩衝することができる貯水池に広がっている。 スライム腺自体はおそらく変更されたcrural腺である。 三つの構造はすべて、他のセグメントの脚対の進化的起源に対応する。
皮膚と筋肉編集
節足動物とは異なり、ベルベットのワームは剛性の外骨格を持っていません。 代わりに、それらの流体で満たされた体腔は、例えばイソギンチャクや様々なワームのような円筒形の多くの無関係な軟体動物と同様に、静水圧骨格とし 体壁上の非圧縮性の内部体液の圧力は剛性を提供し、筋肉はそれに対して作用することができる。 体壁は、非細胞性の外側の皮膚、クチクラで構成されています;内部の皮膚を形成する表皮細胞の単層;そしてこの下に、結合組織に埋め込まれている筋肉の通常3つの層。 クチクラは約マイクロメートルの厚さで、細かい絨毛で覆われています。 組成および構造において、それはα-キチンおよび様々なタンパク質からなる節足動物のクチクラに似ているが、コラーゲンは含まれていない。 それは、外部のepicuticulaと内部のprocuticulaに分けることができ、それ自体はexo-およびendo-cuticulaからなる。 この多重レベル構造はビロードみみずが最も狭い裂け目にそれ自身を絞ることを可能にする外皮の高い柔軟性に責任がある。 外側には撥水性がありますが、クチクラは呼吸による水分の損失を防ぐことができず、その結果、ベルベットの虫は乾燥を避けるために湿度の高い微 クチクラの表面には多数の細かい乳頭が散在しており、そのうちの大きなものは目に見える絨毛様の敏感な毛を運ぶ。 乳頭自体は小さな鱗で覆われており、皮膚にはビロードのような外観があります(そこから一般的な名前が派生している可能性があります)。 それはまた、その撥水性が原因である乾燥したベルベットのように感じます。 皮(ecdysis)の脱皮はホルモンecdysoneによって引き起こされる14日毎にのまわりで、規則的に起こります。 皮膚の内面には六角形の模様があります。 それぞれの脱皮で、小屋の皮膚は、そのすぐ下にある表皮によって置き換えられます。cuticulaとは異なり、これは生きている細胞で構成されています。 これの下にボディの縦方向の軸線に平行か垂直一直線に並ぶコラーゲン繊維で主に構成される結合組織の厚い層があります。 Onychophoraの着色は顔料の範囲によって発生する。 これらの色素の溶解度は有用な診断特性である:すべての節足動物および遅行動物において、体色素はエタノールに可溶性である。 これはPeripatidaeにも当てはまりますが、Peripatopsidaeの場合、体の色素はエタノールに不溶性です。
結合組織の中には、特殊化されていない滑らかな筋肉組織の三つの連続した層があります。 比較的厚い外の層は環状筋肉および縦方向筋肉の同様に大量の内部の層で構成されます。 それらの間には、体の軸に沿って螺旋状に前後に巻く薄い斜めの筋肉があります。 環状および斜めの筋肉の間には、皮膚の表面的に認識可能な横方向のリングの下にあり、擬似セグメント化されたマーキングの原因となる微細な血管 内部筋肉層の下に体腔があります。 断面では、これは、下腹部の中央から上側の縁まで走る、いわゆる背腹筋によって3つの領域に分割される:中央中央部と左右の2つの側部領域には、脚も含まれている。
循環編集
体腔は”pseudocoel”、またはhaemocoelとして知られています。 真の体腔とは異なり、偽体腔は胚中胚葉由来の細胞層によって完全に囲まれていない。 体腔は、しかし、生殖腺と廃棄物を排除する腎症の周りに形成されています。 名前haemocoelが示すように、体腔はすべての器官が埋め込まれる血そっくりの液体で満ちています;このように、それらは血で循環する栄養素と容易に供給す この液体は顔料を含んでいないので無色です; 従って、それは酸素の輸送に於いての限られた役割だけに役立ちます。 アメーバ細胞と腎細胞:血液細胞(または血液細胞)の2つの異なるタイプは、流体中を循環します。 アメーバ細胞は、おそらく細菌や他の異物からの保護に機能し、いくつかの種では、彼らはまた、再生に役割を果たしています。 腎細胞は毒素を吸収するか、またはそれらを腎症による排除に適した形態に変換する。 Haemocoelは、横隔膜によって、背中に沿った心膜洞と腹に沿った心膜洞の2つの部分に分割されます。 前者は管のような心臓を囲み、後者は他の器官を囲む。 ダイヤフラムは多くの場所で穴があい、2つのキャビティ間の液体の交換を可能にする。 心臓自体は、上皮組織からなる環状の筋肉の管であり、セグメントごとに2つの側方開口部(オスティア)を有する。 後端が開いているか閉じているかは分かっていませんが、正面からは体腔内に直接開きます。 血管がないので、体壁の筋肉層とアンテナを供給する一対の動脈の間を走る微細な血管とは別に、これは開放循環と呼ばれます。 ポンプ手順のタイミングは、拡張期と収縮期の2つの部分に分けることができます。 拡張期の間、血液は心膜洞(心臓を含む空洞)からオスティアを通って心臓に流れる。 収縮期が始まると、オスティアが閉じ、心臓の筋肉が内側に収縮し、心臓の容積が減少する。 これは器官を含んでいるperivisceral洞に中心の前端からの血をポンプでくむ。 このようにして、血液が最終的に横隔膜の穿孔を介して心膜洞に戻る前に、様々な器官に栄養素が供給される。 心臓のポンプ作用に加えて、体の動きも循環に影響を与えます。
RespirationEdit
酸素取り込みは、体表面全体を通る単純な拡散を介してある程度起こり、脚のcoxal小胞はいくつかの種に関与している可能性がある。 しかし、最も重要なのは、微細な分岐していない管、気管を介したガス交換であり、これは表面から酸素を様々な器官、特に心臓の深部に引き込む。 これらの構造の壁は、その全体が三マイクロメートル未満の厚さであり、酸素が容易に拡散することができる非常に薄い膜のみで構成されています。 気管は小さな開口部、それ自体が外皮、心房の凹みのような凹部に一緒にクラスタ化されているspiracles、に由来します。 このように形成された”気管束”の数は、体のセグメントごとに平均して約75です; 彼らは生物の背中に最も密集して蓄積します。 節足動物とは異なり、ベルベットのワームは気管の開口部を制御することができません。気管は常に開いており、乾燥した状態ではかなりの水分損失を伴います。 水はミミズの2倍の速さで失われ、毛虫の40倍の速さで失われます。 このため、ベルベットのワームは、高い空気湿度の生息地に依存しています。
DigestionEdit
消化管は頭の少し後ろから始まり、口は体の最前端から少し下に横たわっています。 ここでは、獲物は細かい歯の覆いで下顎骨によって機械的に切断することができます。 二つの唾液腺は、共通の導体を介して、前腸の最初の部分を構成する後続の”喉”に排出される。 それらが産生する唾液には、粘液および加水分解酵素が含まれており、口の中および外で消化を開始する。 喉自体は非常に筋肉質であり、部分的に液化した食物を吸収し、前腸の後部を形成する食道を介して中央腸に送り出すのに役立つ。 前腸とは異なり、これはクチクラで裏打ちされていませんが、代わりに他の動物に見られるように顕著なくぼみを示さない上皮組織の単層のみで構成されています。 中央の腸に入ると、食物粒子は、鋭利な粒子による損傷から腸の内層を保護するのに役立つ粘液ベースのペリトロフィー膜で被覆される。 腸上皮はさらに消化酵素を分泌し、放出された栄養素を吸収するが、消化の大部分はすでに外部または口の中で行われている。 消化できない残党は、再びクチクラで裏打ちされ、肛門で開き、後端の近くの下側に位置する後腸、または直腸に到着する。
ほぼすべてのセグメントには、体腔組織に由来する腎症と呼ばれる一対の排泄器官があります。 それぞれはnephridioporeとして知られている最も近い足の基盤の開始にnephridioductと呼出される鞭毛のコンダクターによって、接続される小さい袋から成っています。 袋はhaemocoelomとnephridium間の仕切りを通した血の限外濾過を促進するpodocytesと呼出される特別な細胞によって占められます。 尿液の組成は、栄養分と水の選択的回収と毒物と廃棄物の分離によってネフリジオダクトで変更され、ネフリジオポアを介して外界に排泄されます。 最も重要な窒素の排泄物プロダクトは水不溶解性の尿酸です;これは少しだけ水との固体状態で、排泄することができます。 このいわゆる尿細管排泄モードは、陸上での生活の調整と、水を経済的に扱う必要性を表しています。 頭部の前腎症のペアは唾液腺に二次的に変換され、男性標本の最終セグメントの別のペアは現在、明らかに生殖に役割を果たす腺として機能してい
SensationEdit
スタブ足を含む体全体には、多数の乳頭が散らばっています: 接触に反応する疣状突起は、先端に機械受容性の剛毛を運び、それぞれが下に横たわっているさらなる感覚神経細胞にも接続されている。 口乳頭、粘液腺の出口は、おそらくまた、感覚知覚にいくつかの機能を持っています。 「唇」またはラブラム上の「感覚細胞」として知られている感覚細胞は、化学刺激に応答し、化学受容体として知られています。 これらはまた、ベルベットワームの最も重要な感覚器官であるように見える二つの触角に見られます。
いくつかの(典型的には地下)種を除いて、一つの単純に構築された目(ocellus)は、各アンテナの後ろに、横方向に、頭のすぐ下にあります。 これはキチン質の球レンズ、角膜および網膜から成り、視神経によって頭脳の中心に接続されます。 網膜は多数の色素細胞および光受容体からなり、後者は容易に改変された鞭毛細胞であり、その鞭毛膜はその表面に感光性色素を運ぶ。 Onychophoraの横紋眼は節足動物の中央ocelliと相同であると考えられています; これは節足動物の最後の共通の祖先が唯一の中央ocelliを持っていた可能性があることを示唆しているでしょう。しかし、神経支配は相同性が限られていることを示しています:Onychophoraの目はアンテナの後ろに形成されますが、反対は節足動物にも当てはまります。
ReproductionEdit
男女ともに生殖腺のペアを所有しており、生殖管と呼ばれるチャネルを介して、後腹側に位置する共通の生殖孔である生殖孔に開 生殖腺および生殖管の両方は、真の体腔組織に由来する。
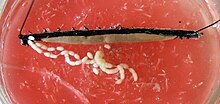
男性は、対応する精子小胞(セミナ小胞)と出口チャネル(vasa efferentia)とともに、二つの別々の精巣を所有しています。 二つのvasa遠心性は、順番にgonoporeで開くために射精チャネルを介して広がる共通の精子管、精管、に団結します。 この横または後ろには、おそらくいくつかの補助生殖機能を果たす特別な腺の2つのペアがあります。 陰茎のような構造は、これまでのところ、Paraperipatus属の男性にのみ見出されているが、まだ作用が観察されていない。
異なる交配手順があります: いくつかの種では、男性は精子を女性の性器の開口部に直接沈着させ、他の種は女性の体に沈着させ、キューティクルが崩壊し、精子細胞が女性に移行することを可能にする。 また、オーストラリアの種では、男性が精子を頭の上に置き、それを女性の性器に押し付けます。 これらの種では頭部に目的が精液を握るおよび/または女性への精液の移動で助けることであるスパイク、棘、空のstylets、ピットおよび不況のような精巧な ほとんどの種の男性はまた女性を引き付けるために足の下側の腺からフェロモンを分泌します。