Les vers de velours sont des animaux segmentés avec une section transversale du corps cylindrique aplatie et des rangées d’appendices corporels non structurés appelés oncopodes ou lobopodes (de manière informelle: pieds en bout). Les animaux atteignent entre 0,5 et 20 cm (.2 à 8 po), la moyenne étant d’environ 5 cm (2 po), et ont entre 13 et 43 paires de pattes. Leur peau se compose de nombreux anneaux transversaux fins et est souvent discrètement colorée en orange, rouge ou marron, mais parfois aussi en vert vif, bleu, or ou blanc, et parfois à motifs d’autres couleurs. La segmentation est extérieurement discrète et identifiable par l’espacement régulier des paires de jambes et par la disposition régulière des pores de la peau, des organes d’excrétion et des concentrations de cellules nerveuses. Les sections individuelles du corps ne sont en grande partie pas spécialisées; même la tête ne se développe que légèrement différemment des segments abdominaux. La segmentation est apparemment spécifiée par le même gène que dans d’autres groupes d’animaux, et est activée dans chaque cas, au cours du développement embryonnaire, à la bordure arrière de chaque segment et dans la zone de croissance des pieds souches. Bien que les onychophores appartiennent au groupe des protostomes, leur développement précoce a une trajectoire deutérostome (la bouche et l’anus se formant séparément); cette trajectoire est dissimulée par les processus assez sophistiqués qui se produisent au début du développement.
Appendices
Les pieds de talon qui caractérisent les vers de velours sont des appendices coniques et amples du corps, qui sont creux à l’intérieur et n’ont pas d’articulations. Bien que le nombre de pieds puisse varier considérablement d’une espèce à l’autre, leur structure est fondamentalement très similaire. La rigidité est assurée par la pression hydrostatique de leur contenu en fluide, et le mouvement est généralement obtenu passivement par étirement et contraction de tout le corps de l’animal. Cependant, chaque jambe peut également être raccourcie et pliée par les muscles internes. En raison du manque d’articulations, cette flexion peut avoir lieu à n’importe quel point sur les côtés de la jambe. Chez certaines espèces, deux organes différents se trouvent dans les pieds:
- Les glandes crurales sont situées à l’épaule des jambes, s’étendant dans la cavité corporelle. Ils s’ouvrent vers l’extérieur au niveau des papilles crurales — de petites bosses ressemblant à des verrues sur le ventre de la jambe – et sécrètent des matériaux messagers chimiques appelés phéromones. Leur nom vient du latin cruralis qui signifie « des jambes ».
- Les vésicules coxales sont des poches situées sur le côté ventre de la jambe, qui peuvent être éventrées et servent probablement à l’absorption d’eau. Ils appartiennent à la famille des Peripatidae et sont nommés de coxa, le mot latin pour « hanche ».

Sur chaque pied se trouve une paire de griffes de chitine rétractables et durcies (sclérotisées), qui donnent la taxon son nom scientifique: Onychophora est dérivé du grec onyches, « griffes »; et phéréine, « porter ». À la base des griffes se trouvent trois à six « coussins » épineux sur lesquels la jambe est assise en position de repos et sur lesquels l’animal marche sur des substrats lisses. Les griffes sont principalement utilisées pour prendre pied fermement sur un terrain accidenté. Chaque griffe est composée de trois éléments empilés, comme des poupées gigognes russes. Le plus externe est versé pendant l’ecdysis, ce qui expose l’élément suivant dans — qui est entièrement formé, n’a donc pas besoin de temps pour durcir avant d’être utilisé. (Cette construction distinctive identifie de nombreux fossiles du Cambrien inférieur comme des ramifications précoces de la lignée des onychophores.) Outre les paires de pattes, il y a trois autres appendices corporels, qui se trouvent à la tête et comprennent trois segments:
- Sur le premier segment de tête se trouve une paire d’antennes minces, qui servent à la perception sensorielle. Ils ne correspondent probablement pas directement aux antennes des Arthropodes, mais peut-être plutôt avec leurs « lèvres » ou leur labrum. À leur base se trouve une paire d’yeux simples, sauf chez quelques espèces aveugles. Devant ceux-ci, chez de nombreuses espèces australiennes, se trouvent diverses fossettes dont la fonction n’est pas encore claire. Il semble que chez certaines espèces au moins, celles-ci servent au transfert de paquets de spermatozoïdes (spermatophores).
- Sur la face ventrale du deuxième segment de la tête se trouve le labrum, une ouverture buccale entourée de « lèvres » sensibles. Chez les vers de velours, cette structure est une excroissance musculaire de la gorge, donc, malgré son nom, elle n’est probablement pas homologue au labrum de l’Arthropode et est utilisée pour l’alimentation. Au fond de la cavité buccale se trouvent les « mâchoires » pointues en forme de croissant, ou mandibules, qui sont fortement durcies et ressemblent aux griffes des pieds, avec lesquelles elles sont homologues en série; au début du développement, les appendices de la mâchoire ont une position et une forme similaires aux jambes suivantes. Les mâchoires sont divisées en mandibules internes et externes et leur surface concave porte de fines denticules. Ils se déplacent en arrière et en avant dans une direction longitudinale, déchirant la proie, apparemment déplacée dans un sens par la musculature et l’autre par la pression hydrostatique. Les griffes sont faites d’α-chitine sclérotisée, renforcées de phénols et de quinones, et ont une composition uniforme – sauf qu’il y a une concentration plus élevée de calcium vers la pointe, ce qui offre probablement une plus grande résistance.
La surface des mandibules est lisse, sans ornementation. La cuticule des mandibules (et des griffes) est distincte du reste du corps. Il a un composant intérieur et extérieur; le composant externe n’a que deux couches (alors que la cuticule du corps en a quatre), et ces couches externes (en particulier l’épicuticule interne) sont déshydratées et fortement tannées, offrant une ténacité.
Glands de slimedit
Sur le troisième segment de tête, à gauche et à droite de la bouche, se trouvent deux ouvertures désignées « papilles buccales ». À l’intérieur de ceux-ci se trouvent une paire de grandes glandes slimes fortement ramifiées à l’intérieur. Ceux-ci se trouvent grossièrement au centre du corps et sécrètent une sorte de boue blanc laiteux, qui est utilisée pour piéger les proies et à des fins défensives. Parfois, le « conducteur de boue » de connexion est élargi dans un réservoir, qui peut amortir la boue pré-produite. Les glandes slimes elles-mêmes sont probablement des glandes crurales modifiées. Les trois structures correspondent à une origine évolutive dans les paires de pattes des autres segments.
Peau et musclemodifier
Contrairement aux arthropodes, les vers de velours ne possèdent pas d’exosquelette rigide. Au lieu de cela, leur cavité corporelle remplie de liquide agit comme un squelette hydrostatique, de la même manière que de nombreux animaux à corps mou non apparentés de forme cylindrique, par exemple des anémones de mer et divers vers. La pression de leur liquide corporel interne incompressible sur la paroi du corps assure la rigidité et les muscles sont capables d’agir contre elle. La paroi du corps se compose d’une peau externe non cellulaire, la cuticula; une seule couche de cellules de l’épiderme formant une peau interne; et en dessous, généralement trois couches de muscles, qui sont intégrées dans les tissus conjonctifs. La cuticula a une épaisseur d’environ un micromètre et est recouverte de fines villosités. Dans sa composition et sa structure, il ressemble à la cuticula des arthropodes, composée d’α-chitine et de diverses protéines, bien que ne contenant pas de collagène. Il peut être divisé en épicuticules externes et procuticules internes, elles-mêmes constituées d’exo- et d’endo-cuticules. Cette structure à plusieurs niveaux est responsable de la grande flexibilité de la peau extérieure, qui permet au ver de velours de se faufiler dans les crevasses les plus étroites. Bien qu’hydrofuge à l’extérieur, la cuticula n’est pas capable d’empêcher la perte d’eau par la respiration et, par conséquent, les vers de velours ne peuvent vivre que dans des microclimats très humides pour éviter la dessiccation. La surface des cuticules est parsemée de nombreuses papilles fines, dont les plus grandes portent des poils sensibles visibles ressemblant à des villosités. Les papilles elles-mêmes sont recouvertes de minuscules écailles, donnant à la peau un aspect velouté (d’où le nom commun est probablement dérivé). Il ressemble également à du velours sec au toucher, dont sa nature hydrofuge est responsable. La mue de la peau (ecdyse) a lieu régulièrement, environ tous les 14 jours, induite par l’hormone ecdysone. La surface interne de la peau porte un motif hexagonal. À chaque mue, la peau perdue est remplacée par l’épiderme, qui se trouve immédiatement en dessous; contrairement à la cuticula, celle-ci est constituée de cellules vivantes. En dessous se trouve une épaisse couche de tissu conjonctif, composée principalement de fibres de collagène alignées parallèlement ou perpendiculairement à l’axe longitudinal du corps. La coloration d’Onychophora est générée par une gamme de pigments. La solubilité de ces pigments est un caractère diagnostique utile: chez tous les arthropodes et tardigrades, le pigment corporel est soluble dans l’éthanol. Cela est également vrai pour les Peripatidae, mais dans le cas des Peripatopsidae, le pigment corporel est insoluble dans l’éthanol.
Dans le tissu conjonctif se trouvent trois couches continues de tissu musculaire lisse non spécialisé. La couche externe relativement épaisse est composée de muscles annulaires et la couche interne tout aussi volumineuse de muscles longitudinaux. Entre eux se trouvent de minces muscles diagonaux qui s’enroulent en arrière et en avant le long de l’axe du corps en spirale. Entre les muscles annulaires et diagonaux existent de fins vaisseaux sanguins, qui se trouvent sous les anneaux transversaux superficiellement reconnaissables de la peau et sont responsables des marques pseudo-segmentées. Sous la couche musculaire interne se trouve la cavité corporelle. En coupe transversale, celle-ci est divisée en trois régions par des muscles dits dorso-ventraux, qui vont du milieu du ventre jusqu’aux bords du côté supérieur: une section médiane centrale et à gauche et à droite, deux régions latérales qui incluent également les jambes.
Circulationmodifier
La cavité corporelle est connue sous le nom de « pseudocoel », ou hémocoel. Contrairement à un véritable cœlome, un pseudocœle n’est pas entièrement entouré d’une couche cellulaire dérivée du mésoderme embryonnaire. Un cœlome se forme cependant autour des gonades et des néphridies qui éliminent les déchets. Comme le suggère le nom haemocoel, la cavité corporelle est remplie d’un liquide semblable à du sang dans lequel tous les organes sont intégrés; de cette manière, ils peuvent être facilement alimentés en nutriments circulant dans le sang. Ce liquide est incolore car il ne contient pas de pigments; pour cette raison, il ne joue qu’un rôle limité dans le transport de l’oxygène. Deux types différents de cellules sanguines (ou hémocytes) circulent dans le liquide: les amibocytes et les néphrocytes. Les amibocytes jouent probablement un rôle de protection contre les bactéries et autres corps étrangers; chez certaines espèces, ils jouent également un rôle dans la reproduction. Les néphrocytes absorbent les toxines ou les convertissent en une forme adaptée à l’élimination par les néphridies. L’hémocèle est divisé par une cloison horizontale, le diaphragme, en deux parties: le sinus péricardique le long du dos et le sinus périviscéral le long du ventre. Le premier renferme le cœur en forme de tube, et le second, les autres organes. Le diaphragme est perforé en de nombreux endroits, permettant l’échange de fluides entre les deux cavités. Le cœur lui-même est un tube de muscles annulaires constitué de tissus épithéliaux, avec deux ouvertures latérales (ostie) par segment. Bien que l’on ne sache pas si l’extrémité arrière est ouverte ou fermée, de l’avant, elle s’ouvre directement dans la cavité corporelle. Comme il n’y a pas de vaisseaux sanguins, à part les vaisseaux fins qui courent entre les couches musculaires de la paroi du corps et une paire d’artères qui alimentent les antennes, on parle de circulation ouverte. Le moment de la procédure de pompage peut être divisé en deux parties: la diastole et la systole. Pendant la diastole, le sang traverse l’ostie du sinus péricardique (la cavité contenant le cœur) dans le cœur. Lorsque la systole commence, l’ostie se ferme et les muscles cardiaques se contractent vers l’intérieur, réduisant ainsi le volume du cœur. Cela pompe le sang de l’extrémité avant du cœur dans le sinus périviscéral contenant les organes. De cette façon, les différents organes sont alimentés en nutriments avant que le sang ne retourne finalement dans le sinus péricardique via les perforations du diaphragme. En plus de l’action de pompage du cœur, les mouvements du corps ont également une influence sur la circulation.
Respirationmodifier
L’absorption d’oxygène se produit dans une certaine mesure par simple diffusion à travers toute la surface du corps, les vésicules coxales sur les pattes pouvant être impliquées chez certaines espèces. Cependant, le plus important est l’échange de gaz via de fins tubes non ramifiés, les trachées, qui aspirent l’oxygène de la surface en profondeur dans les différents organes, en particulier le cœur. Les parois de ces structures, dont l’épaisseur totale est inférieure à trois micromètres, ne sont constituées que d’une membrane extrêmement fine à travers laquelle l’oxygène peut facilement diffuser. Les trachées proviennent de minuscules ouvertures, les spiracles, qui sont eux-mêmes regroupés dans des renfoncements en forme de dent de la peau externe, les oreillettes. Le nombre de » faisceaux de trachées » ainsi formés est en moyenne d’environ 75 par segment corporel; ils s’accumulent le plus densément sur le dos de l’organisme. Contrairement aux arthropodes, les vers de velours sont incapables de contrôler les ouvertures de leurs trachées; les trachées sont toujours ouvertes, entraînant une perte d’eau considérable dans des conditions arides. L’eau est perdue deux fois plus vite que chez les vers de terre et quarante fois plus vite que chez les chenilles. Pour cette raison, les vers de velours dépendent d’habitats très humides.
Digestionmodifier
Le tube digestif commence légèrement derrière la tête, la bouche se trouvant sur la face inférieure un peu à l’avant du corps. Ici, les proies peuvent être démembrées mécaniquement par les mandibules avec leur couverture de fines dents. Deux glandes salivaires se déchargent via un conducteur commun dans la « gorge » suivante, qui constitue la première partie de l’intestin antérieur. La salive qu’ils produisent contient du mucus et des enzymes hydrolytiques, qui initient la digestion dans et à l’extérieur de la bouche. La gorge elle-même est très musclée, servant à absorber les aliments partiellement liquéfiés et à les pomper, via l’œsophage, qui forme la partie arrière de l’intestin antérieur, dans l’intestin central. Contrairement à l’intestin antérieur, celui-ci n’est pas tapissé d’une cuticula, mais se compose uniquement d’une seule couche de tissu épithélial, qui ne présente pas d’indentation visible comme chez d’autres animaux. En entrant dans l’intestin central, les particules alimentaires sont recouvertes d’une membrane péritrophique à base de mucus, qui sert à protéger la muqueuse de l’intestin des dommages causés par des particules à bords tranchants. L’épithélium intestinal sécrète d’autres enzymes digestives et absorbe les nutriments libérés, bien que la majorité de la digestion ait déjà eu lieu à l’extérieur ou dans la bouche. Les restes indigestes arrivent dans l’intestin arrière, ou rectum, qui est à nouveau tapissé d’une cuticula et qui s’ouvre au niveau de l’anus, situé sur la face inférieure près de l’extrémité arrière.
Dans presque tous les segments se trouve une paire d’organes excréteurs appelés néphridies, qui sont dérivés du tissu du coelome. Chacun consiste en une petite poche qui est reliée, via un conducteur flagellé appelé néphridioduct, à une ouverture à la base de la jambe la plus proche connue sous le nom de néphridiopore. La poche est occupée par des cellules spéciales appelées podocytes, qui facilitent l’ultrafiltration du sang à travers la cloison entre l’hémocoélome et le néphridium. La composition de la solution urinaire est modifiée dans le néphridioduc par récupération sélective des nutriments et de l’eau et par isolement du poison et des déchets, avant son excrétion vers le monde extérieur via le néphridiopore. Le produit d’excrétion azoté le plus important est l’acide urique insoluble dans l’eau; il peut être excrété à l’état solide, avec très peu d’eau. Ce mode excréteur uricotélique représente un ajustement à la vie sur terre et la nécessité associée de gérer économiquement l’eau. Une paire d’anciennes néphridies dans la tête a été convertie secondairement en glandes salivaires, tandis qu’une autre paire dans le dernier segment des spécimens mâles sert maintenant de glandes qui jouent apparemment un rôle dans la reproduction.
SensationEdit
Tout le corps, y compris les pieds de talon, est jonché de nombreuses papilles: des protubérances verruqueuses sensibles au toucher qui portent un poil mécanoréceptif à l’extrémité, dont chacune est également connectée à d’autres cellules nerveuses sensorielles situées en dessous. Les papilles buccales, les sorties des glandes slimes, ont probablement également une certaine fonction dans la perception sensorielle. Les cellules sensorielles appelées « sensilles » sur les « lèvres » ou le labrum répondent à des stimuli chimiques et sont connues sous le nom de chimiorécepteurs. Ceux-ci se trouvent également sur les deux antennes, qui semblent être les organes sensoriels les plus importants du ver de velours.
À l’exception de quelques espèces (généralement souterraines), un œil simplement construit (ocellus) se trouve derrière chaque antenne, latéralement, juste en dessous de la tête. Il se compose d’une lentille de boule chitineuse, d’une cornée et d’une rétine et est relié au centre du cerveau par l’intermédiaire d’un nerf optique. La rétine comprend de nombreuses cellules pigmentaires et photorécepteurs ; ces dernières sont des cellules flagellées facilement modifiées, dont les membranes du flagelle portent un pigment photosensible à leur surface. On pense que les yeux rhabdomériques de l’Onychophora sont homologues aux ocelles médians des arthropodes; cela suggère que le dernier ancêtre commun des arthropodes n’avait peut-être que des ocelles médians.Cependant, l’innervation montre que l’homologie est limitée : les yeux d’Onychophora se forment derrière l’antenne, alors que l’inverse est vrai chez les arthropodes.
Reproductionmodifier
Les deux sexes possèdent des paires de gonades qui s’ouvrent par un canal appelé gonoduc dans une ouverture génitale commune, le gonopore, située sur la face ventrale arrière. Les gonades et le gonoduc sont tous deux dérivés du véritable tissu du cœlome.
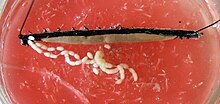
Chez les femelles, les deux ovaires sont joints au milieu et au diaphragme horizontal. Le gonoduc apparaît différemment selon que l’espèce est vivante ou pondeuse. Chez les espèces vivantes, chaque canal de sortie se divise en un oviducte mince et un « utérus » spacieux, l’utérus, dans lequel les embryons se développent. Le vagin unique, auquel les deux utérus sont connectés, s’étend vers l’extérieur jusqu’au gonopore. Chez les espèces pondeuses, dont le gonoduc est uniformément construit, l’ouverture génitale se trouve à l’extrémité d’un long appareil de ponte, l’ovipositeur. Les femelles de nombreuses espèces possèdent également un dépôt de sperme appelé réceptacle seminis, dans lequel les spermatozoïdes des mâles peuvent être stockés temporairement ou pendant de plus longues périodes.
Les mâles possèdent deux testicules séparés, ainsi que la vésicule spermatique correspondante (la vesicula seminalis) et le canal de sortie (la vasa efferentia). Les deux vasa efferentia s’unissent à un canal spermatique commun, le canal déférent, qui à son tour s’élargit à travers le canal éjaculatoire pour s’ouvrir au niveau du gonopore. Juste à côté ou derrière cela se trouvent deux paires de glandes spéciales, qui remplissent probablement une fonction de reproduction auxiliaire; les glandes les plus en arrière sont également appelées glandes anales. Jusqu’à présent, une structure ressemblant à un pénis n’a été trouvée que chez les mâles du genre Paraperipatus, mais n’a pas encore été observée en action.
Il existe différentes procédures d’accouplement: Chez certaines espèces, les mâles déposent leur spermatophore directement dans les organes génitaux de la femelle, tandis que d’autres le déposent sur le corps de la femelle, où la cuticule s’effondre et permet aux spermatozoïdes de migrer dans la femelle. Il existe également des espèces australiennes où le mâle place son spermatophore sur le dessus de sa tête, qui est ensuite pressé contre les organes génitaux de la femelle. Chez ces espèces, la tête a des structures élaborées comme des pointes, des épines, des stylets creux, des fosses et des dépressions, dont le but est de retenir le sperme et / ou d’aider au transfert du sperme à la femelle. Les mâles de la plupart des espèces sécrètent également une phéromone des glandes situées sous les pattes pour attirer les femelles.