I vermi di velluto sono animali segmentati con una sezione cilindrica appiattita del corpo e file di appendici del corpo non strutturate note come oncopodi o lobopodi (informalmente: piedi stub). Gli animali crescono tra 0,5 e 20 cm (.2 a 8 in), con la media di circa 5 cm (2 in), e hanno tra 13 e 43 paia di zampe. La loro pelle è costituita da numerosi, sottili anelli trasversali ed è spesso poco appariscente di colore arancione, rosso o marrone, ma a volte anche verde brillante, blu, oro o bianco, e occasionalmente modellata con altri colori. La segmentazione è esteriormente poco appariscente e identificabile dalla spaziatura regolare delle coppie di gambe e dalla disposizione regolare dei pori della pelle, degli organi di escrezione e delle concentrazioni delle cellule nervose. Le singole sezioni del corpo sono in gran parte non specializzate; anche la testa si sviluppa in modo leggermente diverso dai segmenti addominali. La segmentazione è apparentemente specificata dallo stesso gene di altri gruppi di animali e viene attivata in ciascun caso, durante lo sviluppo embrionale, al bordo posteriore di ciascun segmento e nella zona di crescita dei piedi del mozzicone. Sebbene gli onicoforani rientrino nel gruppo dei protostomi, il loro sviluppo precoce ha una traiettoria deuterostomica, (con la bocca e l’ano che si formano separatamente); questa traiettoria è nascosta dai processi piuttosto sofisticati che si verificano nello sviluppo precoce.
appendiceedit
I piedi mozzati che caratterizzano i vermi di velluto sono appendici coniche e larghe del corpo, che sono internamente cave e prive di giunture. Anche se il numero di piedi può variare notevolmente tra le specie, la loro struttura è fondamentalmente molto simile. La rigidità è fornita dalla pressione idrostatica del loro contenuto fluido e il movimento è solitamente ottenuto passivamente allungando e riducendo l’intero corpo dell’animale. Tuttavia, ogni gamba può anche essere accorciata e piegata dai muscoli interni. A causa della mancanza di articolazioni, questa piegatura può avvenire in qualsiasi punto lungo i lati della gamba. In alcune specie, due diversi organi si trovano all’interno dei piedi:
- Le ghiandole crurali si trovano alla spalla delle gambe, estendendosi nella cavità del corpo. Si aprono verso l’esterno alle papille crurali—piccole protuberanze simili a verruche sul lato della pancia della gamba-e secernono materiali messaggeri chimici chiamati feromoni. Il loro nome deriva dal latino cruralis che significa “delle gambe”.
- Le vescicole di Coxal sono sacchetti situati sul lato della pancia della gamba, che possono essere everted e probabilmente servire nell’assorbimento di acqua. Appartengono alla famiglia Peripatidae e prendono il nome da coxa, la parola latina per “anca”.

Su ogni piede è una coppia di retrattile, temprato (sclerotised) chitina artigli, che danno il taxon il suo nome scientifico: Onychophora deriva dal greco onyches, “artigli”; e pherein, di “portare”. Alla base degli artigli ci sono da tre a sei “cuscini” spinosi su cui la gamba si siede nella sua posizione di riposo e su cui l’animale cammina su substrati lisci. Gli artigli sono usati principalmente per ottenere un punto d’appoggio fermo su terreni sconnessi. Ogni artiglio è composto da tre elementi impilati, come bambole russe nidificanti. Il più esterno viene versato durante l’ecdisi, che espone l’elemento successivo in — che è completamente formato, quindi non ha bisogno di tempo per indurirsi prima di essere utilizzato. (Questa costruzione distintiva identifica molti fossili del Cambriano come prime propaggini del lignaggio onicoforo.) Oltre alle paia di zampe, ci sono tre ulteriori appendici del corpo, che sono alla testa e comprendono tre segmenti:
- Sul primo segmento della testa c’è una coppia di antenne snelle, che servono nella percezione sensoriale. Probabilmente non corrispondono direttamente alle antenne degli Artropodi, ma forse piuttosto con le loro “labbra” o labrum. Alla loro base si trova un paio di occhi semplici, tranne che in alcune specie cieche. Di fronte a questi, in molte specie australiane, ci sono varie fossette, la cui funzione non è ancora chiara. Sembra che in almeno alcune specie, questi servano nel trasferimento di pacchetti di cellule spermatiche (spermatofori).
- Sul lato della pancia del secondo segmento di testa si trova il labrum, un’apertura della bocca circondata da “labbra”sensibili. Nei vermi di velluto, questa struttura è una crescita muscolare della gola, quindi, nonostante il suo nome, probabilmente non è omologa al labrum dell’Artropoda e viene utilizzata per l’alimentazione. In profondità all’interno della cavità orale si trovano le “mascelle” affilate a forma di mezzaluna, o mandibole, che sono fortemente indurite e assomigliano agli artigli dei piedi, con i quali sono omologhi in serie; all’inizio dello sviluppo, le appendici della mascella hanno una posizione e una forma simili alle gambe successive. Le mascelle sono divise in mandibole interne ed esterne e la loro superficie concava reca dentelli fini. Si muovono avanti e indietro in direzione longitudinale, strappando la preda, apparentemente mossa in una direzione dalla muscolatura e l’altra dalla pressione idrostatica. Gli artigli sono fatti di α-chitina sclerotizzata, rinforzata con fenoli e chinoni, e hanno una composizione uniforme – tranne che c’è una maggiore concentrazione di calcio verso la punta, presumibilmente offrendo una maggiore resistenza.
La superficie delle mandibole è liscia, senza ornamenti. La cuticola delle mandibole (e degli artigli) è distinta dal resto del corpo. Ha un componente interno ed esterno; il componente esterno ha solo due strati (mentre la cuticola del corpo ne ha quattro) e questi strati esterni (in particolare l’epicuticolo interno) sono disidratati e fortemente abbronzati, offrendo tenacità.
Slime glandsEdit
Sul terzo segmento di testa, a sinistra ea destra della bocca, ci sono due aperture designate “papille orali”. All’interno di questi ci sono un paio di grandi ghiandole melmose fortemente ramificate internamente. Questi si trovano approssimativamente al centro del corpo e secernono una sorta di melma bianco latte, che viene utilizzata per intrappolare le prede e per scopi difensivi. A volte il “conduttore di melma” di collegamento viene ampliato in un serbatoio, che può tamponare la melma pre-prodotta. Le ghiandole melmose stesse sono probabilmente ghiandole crurali modificate. Tutte e tre le strutture corrispondono a un’origine evolutiva nelle coppie di gambe degli altri segmenti.
Pelle e muscolomodifica
A differenza degli artropodi, i vermi di velluto non possiedono un esoscheletro rigido. Invece, la loro cavità corporea piena di liquido agisce come uno scheletro idrostatico, in modo simile a molti animali dal corpo morbido non correlati che sono di forma cilindrica, ad esempio anemoni di mare e vari vermi. La pressione del loro fluido corporeo interno incomprimibile sulla parete del corpo fornisce rigidità e i muscoli sono in grado di agire contro di esso. La parete del corpo è costituita da una pelle esterna non cellulare, la cuticola; un singolo strato di cellule epidermiche che formano una pelle interna; e sotto questo, di solito tre strati di muscoli, che sono incorporati nei tessuti connettivi. La cuticola è spessa circa un micrometro e ricoperta di villi fini. Nella composizione e nella struttura, assomiglia alla cuticola degli artropodi, costituita da α-chitina e varie proteine, sebbene non contenga collagene. Può essere diviso in un epicuticolo esterno e un procuticolo interno, che a loro volta consistono in eso – ed cu-cuticola. Questa struttura multilivello è responsabile dell’elevata flessibilità della pelle esterna, che consente al verme di velluto di spremersi nelle fessure più strette. Sebbene esteriormente idrorepellente, la cuticola non è in grado di prevenire la perdita di acqua dalla respirazione e, di conseguenza, i vermi di velluto possono vivere solo in microclimi con elevata umidità per evitare l’essiccamento. La superficie della cuticola è disseminata di numerose papille fini, le più grandi delle quali portano setole sensibili simili a villi visibili. Le papille stesse sono ricoperte da minuscole squame, conferendo alla pelle un aspetto vellutato (da cui probabilmente deriva il nome comune). Sembra anche un velluto secco al tatto, di cui è responsabile la sua natura idrorepellente. La muta della pelle (ecdisi) avviene regolarmente, circa ogni 14 giorni, indotta dall’ormone ecdysone. La superficie interna della pelle ha un motivo esagonale. Ad ogni muta, la pelle del capannone viene sostituita dall’epidermide, che si trova immediatamente sotto di essa; a differenza della cuticola, questa consiste di cellule viventi. Sotto questo si trova uno spesso strato di tessuto connettivo, che è composto principalmente da fibre di collagene allineate parallele o perpendicolari all’asse longitudinale del corpo. La colorazione di Onychophora è generata da una gamma di pigmenti. La solubilità di questi pigmenti è un utile carattere diagnostico: in tutti gli artropodi e tardigradi, il pigmento corporeo è solubile in etanolo. Questo vale anche per i Peripatidae, ma nel caso dei Peripatopsidae, il pigmento del corpo è insolubile in etanolo.
All’interno del tessuto connettivo si trovano tre strati continui di tessuto muscolare liscio non specializzato. Lo strato esterno relativamente spesso è composto da muscoli anulari e lo strato interno altrettanto voluminoso di muscoli longitudinali. Tra di loro si trovano sottili muscoli diagonali che si snodano indietro e in avanti lungo l’asse del corpo in una spirale. Tra i muscoli anulari e diagonali esistono vasi sanguigni sottili, che si trovano al di sotto degli anelli trasversali superficialmente riconoscibili della pelle e sono responsabili delle marcature pseudo-segmentate. Sotto lo strato muscolare interno si trova la cavità del corpo. Nella sezione trasversale, questo è diviso in tre regioni dai cosiddetti muscoli dorso-ventrali, che corrono dal centro del ventre fino ai bordi del lato superiore: una sezione centrale e, a sinistra ea destra, due regioni laterali che includono anche le gambe.
CirculationEdit
La cavità corporea è nota come “pseudocoel”, o haemocoel. A differenza di un vero celoma, uno pseudocoel non è completamente racchiuso da uno strato cellulare derivato dal mesoderma embrionale. Un celoma è, tuttavia, formato intorno alle gonadi e ai nefridi che eliminano i rifiuti. Come suggerisce il nome haemocoel, la cavità del corpo è riempita con un liquido simile al sangue in cui sono incorporati tutti gli organi; in questo modo, possono essere facilmente forniti con sostanze nutritive che circolano nel sangue. Questo liquido è incolore in quanto non contiene pigmenti; per questo motivo, serve solo un ruolo limitato nel trasporto di ossigeno. Due diversi tipi di cellule del sangue (o emociti) circolano nel fluido: amebociti e nefrociti. Gli amebociti probabilmente funzionano in protezione da batteri e altri corpi estranei; in alcune specie, svolgono anche un ruolo nella riproduzione. I nefrociti assorbono le tossine o le convertono in una forma adatta all’eliminazione da parte dei nefridi. L’emocoel è diviso da una partizione orizzontale, il diaframma, in due parti: il seno pericardico lungo la schiena e il seno periviscerale lungo la pancia. Il primo racchiude il cuore a forma di tubo, e il secondo, gli altri organi. Il diaframma è perforato in molti punti, consentendo lo scambio di fluidi tra le due cavità. Il cuore stesso è un tubo di muscoli anulari costituito da tessuti epiteliali, con due aperture laterali (ostia) per segmento. Mentre non è noto se l’estremità posteriore è aperta o chiusa, dalla parte anteriore, si apre direttamente nella cavità del corpo. Poiché non ci sono vasi sanguigni, a parte i vasi sottili che corrono tra gli strati muscolari della parete del corpo e una coppia di arterie che forniscono le antenne, questo è indicato come una circolazione aperta. I tempi della procedura di pompaggio possono essere suddivisi in due parti: diastole e sistole. Durante la diastole, il sangue scorre attraverso l’ostia dal seno pericardico (la cavità contenente il cuore) nel cuore. Quando inizia la sistole, l’ostia si chiude e i muscoli cardiaci si contraggono verso l’interno, riducendo il volume del cuore. Questo pompa il sangue dall’estremità anteriore del cuore nel seno periviscerale contenente gli organi. In questo modo, i vari organi vengono forniti con sostanze nutritive prima che il sangue ritorni finalmente al seno pericardico attraverso le perforazioni nel diaframma. Oltre all’azione di pompaggio del cuore, i movimenti del corpo hanno anche un’influenza sulla circolazione.
Respirazionemodifica
L’assorbimento di ossigeno si verifica in una certa misura attraverso la semplice diffusione attraverso l’intera superficie corporea, con le vescicole coxali sulle gambe che possono essere coinvolte in alcune specie. Tuttavia, di maggior importanza è lo scambio di gas attraverso sottili tubi non ramificati, le trachee, che attingono ossigeno dalla superficie in profondità nei vari organi, in particolare il cuore. Le pareti di queste strutture, che sono spesse meno di tre micrometri nella loro interezza, consistono solo di una membrana estremamente sottile attraverso la quale l’ossigeno può facilmente diffondersi. Le trachee hanno origine in minuscole aperture, gli spiracoli, che a loro volta sono raggruppati in recessi simili a denti della pelle esterna, gli atri. Il numero di” fasci di trachee ” così formati è in media intorno a 75 per segmento corporeo; si accumulano più densamente sul retro dell’organismo. A differenza degli artropodi, i vermi di velluto non sono in grado di controllare le aperture delle loro trachee; le trachee sono sempre aperte, comportando una notevole perdita d’acqua in condizioni aride. L’acqua viene persa due volte più velocemente dei lombrichi e quaranta volte più velocemente rispetto ai bruchi. Per questo motivo, i vermi di velluto dipendono da habitat con elevata umidità dell’aria.
Digestionemodifica
Il tratto digestivo inizia leggermente dietro la testa, la bocca giace sul lato inferiore un po ‘ dal punto più frontale del corpo. Qui, le prede possono essere smembrate meccanicamente dalle mandibole con la loro copertura di denti sottili. Due ghiandole salivari scaricano attraverso un conduttore comune nella successiva “gola”, che costituisce la prima parte dell’intestino anteriore. La saliva che producono contiene muco ed enzimi idrolitici, che iniziano la digestione dentro e fuori la bocca. La gola stessa è molto muscolosa, serve ad assorbire il cibo parzialmente liquefatto e a pomparlo, tramite l’esofago, che costituisce la parte posteriore dell’intestino anteriore, nell’intestino centrale. A differenza dell’intestino anteriore, questo non è rivestito da una cuticola, ma consiste solo di un singolo strato di tessuto epiteliale, che non presenta una rientranza cospicua come si trova in altri animali. Entrando nell’intestino centrale, le particelle di cibo sono rivestite con una membrana peritrofica a base di muco, che serve a proteggere il rivestimento dell’intestino dai danni causati da particelle taglienti. L’epitelio intestinale secerne ulteriori enzimi digestivi e assorbe i nutrienti rilasciati, sebbene la maggior parte della digestione sia già avvenuta esternamente o in bocca. I resti indigeribili arrivano nell’intestino posteriore, o retto, che è ancora una volta rivestito da una cuticola e che si apre all’ano, situato sul lato inferiore vicino all’estremità posteriore.
In quasi ogni segmento è una coppia di organi escretori chiamati nefridi, che sono derivati dal tessuto celom. Ciascuno è costituito da una piccola sacca che è collegata, tramite un conduttore flagellato chiamato nefridiodotto, ad un’apertura alla base della gamba più vicina nota come nefridioporo. La sacca è occupata da cellule speciali chiamate podociti, che facilitano l’ultrafiltrazione del sangue attraverso la partizione tra emocoelom e nefridio. La composizione della soluzione urinaria viene modificata nel nefridiodotto dal recupero selettivo di nutrienti e acqua e dall’isolamento di veleno e materiali di scarto, prima che venga escreta nel mondo esterno attraverso il nefridioporo. Il più importante prodotto di escrezione azotata è l’acido urico insolubile in acqua; questo può essere escreto allo stato solido, con pochissima acqua. Questa cosiddetta modalità escretoria uricotelica rappresenta un adattamento alla vita sulla terra e la necessità associata di trattare economicamente con l’acqua. Un paio di precedenti nefridi nella testa sono stati convertiti secondariamente nelle ghiandole salivari, mentre un altro paio nel segmento finale di esemplari maschi ora servono come ghiandole che apparentemente svolgono un ruolo nella riproduzione.
SensationEdit
L’intero corpo, compresi i piedi del mozzicone, è disseminato di numerose papille: sporgenze verrucose sensibili al tatto che portano una setola meccanorecettiva sulla punta, ognuna delle quali è anche collegata a ulteriori cellule nervose sensoriali che si trovano sotto. Le papille della bocca, le uscite delle ghiandole melma, probabilmente hanno anche una certa funzione nella percezione sensoriale. Le cellule sensoriali conosciute come” sensills “sulle” labbra ” o labrum rispondono agli stimoli chimici e sono conosciute come chemorecettori. Questi si trovano anche sulle due antenne, che sembrano essere gli organi sensoriali più importanti del verme di velluto.
Tranne che in alcune specie (tipicamente sotterranee), un occhio semplicemente costruito (ocellus) si trova dietro ogni antenna, lateralmente, appena sotto la testa. Questo è costituito da una lente a sfera chitinosa, una cornea e una retina ed è collegato al centro del cervello tramite un nervo ottico. La retina comprende numerose cellule pigmentate e fotorecettori; questi ultimi sono cellule flagellate facilmente modificate, le cui membrane flagellali portano un pigmento fotosensibile sulla loro superficie. Gli occhi rabdomerici degli Onychophora sono ritenuti omologhi con gli ocelli mediani degli artropodi; ciò suggerirebbe che l’ultimo antenato comune degli artropodi potesse avere solo ocelli mediani.Tuttavia, l’innervazione mostra che l’omologia è limitata: gli occhi di Onychophora si formano dietro l’antenna, mentre è vero il contrario negli artropodi.
RiproduzioneEdit
Entrambi i sessi possiedono coppie di gonadi, che si aprono attraverso un canale chiamato gonodotto in un’apertura genitale comune, il gonoporo, che si trova sul lato ventrale posteriore. Sia le gonadi che il gonodotto derivano dal vero tessuto del celoma.
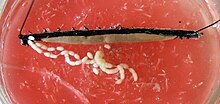
Nelle femmine, le due ovaie sono unite al centro e al diaframma orizzontale. Il gonodotto appare in modo diverso a seconda che la specie sia viva o ovodeposta. Nelle specie viventi, ogni canale di uscita si divide in un ovidotto snello e un “utero” spazioso, l’utero, in cui si sviluppano gli embrioni. La singola vagina, a cui sono collegati entrambi gli uteri, corre verso l’esterno verso il gonoporo. Nelle specie ovaiole, il cui gonodotto è costruito in modo uniforme, l’apertura genitale si trova sulla punta di un lungo apparato ovopositore, l’ovopositore. Le femmine di molte specie possiedono anche un deposito di sperma chiamato recipiente seminis, in cui le cellule spermatiche dei maschi possono essere conservate temporaneamente o per periodi più lunghi.
I maschi possiedono due testicoli separati, insieme alla corrispondente vescicola spermatica (la vescicola seminalis) e al canale di uscita (la vasa efferentia). I due vasa efferentia si uniscono ad un dotto spermatico comune, il dotto deferente, che a sua volta si allarga attraverso il canale eiaculatorio per aprirsi al gonoporo. Direttamente accanto o dietro questa si trovano due paia di ghiandole speciali, che probabilmente servono qualche funzione riproduttiva ausiliaria; le ghiandole più arretrate sono anche conosciute come ghiandole anali. Una struttura simile al pene è stata finora trovata solo nei maschi del genere Paraperipatus ma non è stata ancora osservata in azione.
Esistono diverse procedure di accoppiamento: In alcune specie i maschi depositano il loro spermatoforo direttamente nell’apertura dei genitali della femmina, mentre altri lo depositano sul corpo della femmina, dove la cuticola collasserà e permettendo alle cellule spermatiche di migrare nella femmina. Ci sono anche specie australiane in cui il maschio posiziona il loro spermatoforo sopra la testa, che viene poi premuto contro i genitali della femmina. In queste specie la testa ha strutture elaborate come punte, spine, stiletti cavi, fosse e depressioni, il cui scopo è quello di trattenere lo sperma e / o assistere nel trasferimento dello sperma alla femmina. I maschi della maggior parte delle specie secernono anche un feromone dalle ghiandole sul lato inferiore delle gambe per attirare le femmine.