fluweelwormen zijn gesegmenteerde dieren met een afgeplatte cilindrische dwarsdoorsnede van het lichaam en Rijen ongestructureerde aanhangsels die bekend staan als oncopoden of lobopoden (informeel: stubpoten). De dieren worden tussen de 0,5 en 20 cm (.2 tot 8 in), met een gemiddelde van ongeveer 5 cm (2 in), en hebben tussen 13 en 43 paar poten. Hun huid bestaat uit talrijke, fijne dwarsringen en is vaak onopvallend oranje, rood of bruin gekleurd, maar soms ook heldergroen, blauw, goud of wit, en soms voorzien van andere kleuren. De segmentatie is uiterlijk onopvallend, en herkenbaar door de regelmatige afstand tussen de paar benen en in de regelmatige opstelling van huidporiën, uitscheidingsorganen en concentraties van zenuwcellen. De individuele lichaamsdelen zijn grotendeels ongespecificeerd; zelfs het hoofd ontwikkelt zich slechts een beetje anders dan de abdominale segmenten. Segmentatie wordt blijkbaar gespecificeerd door hetzelfde gen als bij andere groepen dieren, en wordt in elk geval geactiveerd, tijdens de embryonale ontwikkeling, aan de achterste grens van elk segment en in de groeizone van de stubvoeten. Hoewel onychophoranen binnen de protostoomgroep vallen, heeft hun vroege ontwikkeling een deuterostoombaan (waarbij de mond en de anus zich afzonderlijk vormen); dit traject wordt verborgen door de vrij verfijnde processen die in vroege ontwikkeling voorkomen.
aanhangsel Edit
De stubpoten die de fluweelwormen kenmerken zijn conische, baggy aanhangsels van het lichaam, die inwendig hol zijn en geen gewrichten hebben. Hoewel het aantal voeten aanzienlijk kan variëren tussen soorten, is hun structuur in principe zeer vergelijkbaar. Stijfheid wordt geboden door de hydrostatische druk van hun vloeistofinhoud, en beweging wordt meestal passief verkregen door uitrekken en samentrekking van het hele lichaam van het dier. Echter, elk been kan ook worden ingekort en gebogen door interne spieren. Door het gebrek aan gewrichten kan deze buiging plaatsvinden op elk punt langs de zijkanten van het been. Bij sommige soorten worden twee verschillende organen in de voeten aangetroffen:
- Keurklieren bevinden zich aan de schouder van de benen, tot in de lichaamsholte. Ze openen naar buiten bij de crural papillen-kleine wrat-achtige bulten op de buikzijde van het been—en scheiden chemische boodschapper materialen genaamd feromonen. Hun naam komt van het Latijnse cruralis wat “van de benen” betekent.
- Coxale blaasjes zijn buidels aan de buikzijde van het been, die kunnen worden doorboord en waarschijnlijk dienen voor waterabsorptie. Ze behoren tot de familie Peripatidae en zijn vernoemd naar coxa, het Latijnse woord voor “hip”.

Op elke voet is een paar intrekbare, gehard (sclerotised) chitine klauwen, die geven het taxon zijn wetenschappelijke naam: Onychophora is afgeleid van het griekse onyches, “klauwen”; en pherein, “dragen”. Aan de basis van de klauwen bevinden zich drie tot zes stekelige “kussens” waarop het been in zijn rustpositie zit en waarop het dier over gladde ondergronden loopt. De klauwen worden vooral gebruikt om een stevige voet aan de grond te krijgen op oneffen terrein. Elke klauw bestaat uit drie gestapelde elementen, zoals Russische nestpoppen. De buitenste wordt vergoten tijdens ecdysis, die bloot het volgende element in — die volledig is gevormd, dus geen tijd nodig om uit te harden voordat het wordt gebruikt. (Deze kenmerkende constructie identificeert veel vroege Cambrische fossielen als vroege uitlopers van de onychophoraanse afstamming.) Naast de paar poten zijn er nog drie andere lichaamsaanhangsels, die zich aan het hoofd bevinden en drie segmenten omvatten:
- op het eerste kopsegment bevindt zich een paar slanke antennes, die dienen voor de zintuiglijke waarneming. Ze komen waarschijnlijk niet direct overeen met de antennes van de Arthropoda, maar misschien eerder met hun “lippen” of labrum. Aan de basis is een paar eenvoudige ogen te vinden, behalve bij een paar blinde soorten. Voor deze, bij veel Australische soorten, zijn verschillende kuiltjes, waarvan de functie nog niet duidelijk is. Het blijkt dat deze in ten minste sommige soorten dienen bij de overdracht van zaadcelpakketten (spermatoforen).
- op de buikzijde van het tweede hoofdsegment bevindt zich het labrum, een mondopening omgeven door gevoelige “lippen”. In de fluwelen wormen is deze structuur een spieruitgroei van de keel, dus, ondanks de naam, is het waarschijnlijk niet homoloog aan het labrum van de Arthropoda en wordt gebruikt voor het voeden. Diep in de mondholte liggen de scherpe, halvemaanvormige “kaken”, of mandibels, die sterk gehard zijn en lijken op de klauwen van de voeten, waarmee ze serieel homoloog zijn; vroeg in ontwikkeling, de kaak aanhangsels hebben een positie en vorm vergelijkbaar met de volgende benen. De kaken zijn verdeeld in interne en externe mandibels en hun concave oppervlak draagt fijne denticles. Ze bewegen naar achteren en naar voren in een longitudinale richting, scheuren de prooi uit elkaar, blijkbaar verplaatst in de ene richting door spiermassa en de andere door hydrostatische druk. De klauwen zijn gemaakt van gesclerotiseerd α-chitine, versterkt met fenolen en chinonen, en hebben een uniforme samenstelling – behalve dat er een hogere concentratie van calcium naar de top is, vermoedelijk met een grotere sterkte.
het oppervlak van de mandibels is glad, zonder versiering. De cuticula in de mandibels (en klauwen) is te onderscheiden van de rest van het lichaam. Het heeft een binnen-en buitencomponent; de buitenste component heeft slechts twee lagen (terwijl het lichaam cuticula heeft vier), en deze buitenste lagen (in het bijzonder de binnenste epicuticula) zijn uitgedroogd en sterk gebruind, waardoor taaiheid.
Slime glandsEdit
op het derde kopsegment, links en rechts van de mond, zijn twee openingen aangeduid als “orale papillen”. Daarin bevinden zich een paar grote, sterk inwendig vertakte slijmklieren. Deze liggen ruwweg in het midden van het lichaam en scheiden een soort melkwit slijm af, dat wordt gebruikt om prooi te verstrikken en voor defensieve doeleinden. Soms wordt de verbindende “slijmgeleider” verbreed tot een reservoir, dat vooraf geproduceerd slijm kan bufferen. De slijmklieren zelf zijn waarschijnlijk gemodificeerde crural klieren. Alle drie de structuren komen overeen met een evolutionaire oorsprong in de beenparen van de andere segmenten.
huid en muscleEdit
in tegenstelling tot geleedpotigen hebben fluweelwormen geen stijf exoskelet. In plaats daarvan, hun met vloeistof gevulde lichaamsholte fungeert als een hydrostatisch skelet, net als veel ongerelateerde zacht-bodied dieren die cilindrisch gevormd zijn, bijvoorbeeld zeeanemonen en verschillende wormen. Druk van hun incompressible interne lichaamsvocht op de lichaamswand zorgt voor stijfheid, en spieren zijn in staat om tegen te handelen. De wand van het lichaam bestaat uit een niet-cellulaire buitenhuid, de cuticula; een enkele laag van epidermiscellen vormen een interne huid; en daaronder, meestal drie lagen van spier, die zijn ingebed in bindweefsels. De cuticula is ongeveer een micrometer dik en bedekt met fijne villi. Qua samenstelling en structuur lijkt het op de cuticula van de geleedpotigen, bestaande uit α-chitine en verschillende eiwitten, hoewel het geen collageen bevat. Het kan worden onderverdeeld in een externe epicuticula en een interne prouticula, die zelf bestaan uit exo – en endo-cuticula. Deze multi-level structuur is verantwoordelijk voor de hoge flexibiliteit van de buitenhuid, waardoor de fluweelworm zich in de smalste spleten kan persen. Hoewel de cuticula naar buiten waterafstotend is, is het niet in staat om waterverlies door ademhaling te voorkomen, en als gevolg daarvan kunnen fluweelwormen alleen in microklimaten met een hoge luchtvochtigheid leven om uitdroging te voorkomen. Het oppervlak van de cuticula is bezaaid met tal van fijne papillen, waarvan de grootste zichtbare villi-achtige gevoelige haren dragen. De papillen zelf zijn bedekt met kleine schubben, waardoor de huid een fluweelzacht uiterlijk heeft (waarvan de gewone naam waarschijnlijk is afgeleid). Het voelt ook als droog fluweel aan, waarvoor de waterafstotende aard verantwoordelijk is. Rui van de huid (ecdysis) vindt regelmatig plaats, ongeveer om de 14 dagen, veroorzaakt door het hormoon ecdyson. Het binnenoppervlak van de huid draagt een zeshoekig patroon. Bij elke rui wordt de afgeworpen huid vervangen door de epidermis, die er direct onder ligt; in tegenstelling tot de cuticula bestaat deze uit levende cellen. Daaronder ligt een dikke laag bindweefsel, die voornamelijk bestaat uit collageenvezels die evenwijdig of loodrecht op de lengteas van het lichaam zijn uitgelijnd. De kleuring van Onychophora wordt gegenereerd door een reeks pigmenten. De oplosbaarheid van deze pigmenten is een nuttig Diagnostisch karakter: in alle geleedpotigen en tardigrades is het lichaamspigment oplosbaar in ethanol. Dit geldt ook voor de Peripatidae, maar bij de Peripatopsidae is het lichaamspigment onoplosbaar in ethanol.
binnen het bindweefsel liggen drie continue lagen van niet-gespecificeerd gladde spierweefsel. De relatief dikke buitenlaag bestaat uit ringvormige spieren en de evenzo volumineuze binnenlaag van longitudinale spieren. Daartussen liggen dunne diagonale spieren die in een spiraal naar achteren en naar voren kronkelen langs de lichaams-as. Tussen de ringvormige en diagonale spieren bestaan fijne bloedvaten, die onder de oppervlakkig herkenbare dwarsringen van de huid liggen en verantwoordelijk zijn voor de pseudo-gesegmenteerde markeringen. Onder de inwendige spierlaag ligt de lichaamsholte. In doorsnede is dit verdeeld in drie gebieden door zogenaamde dorso-ventrale spieren, die lopen van het midden van de onderbuik door naar de randen van de bovenzijde: een centrale buik en links en rechts, twee zijdelingse gebieden die ook de benen.
Circulatiedit
de lichaamsholte staat bekend als een “pseudokoel” of hemocoel. In tegenstelling tot een echte coelom, is een pseudocoel niet volledig ingesloten door een cellaag afgeleid van het embryonale mesoderm. Een coelom wordt echter gevormd rond de geslachtsklieren en de afvalverwijdende nefridia. Zoals de naam hemocoel suggereert, is de lichaamsholte gevuld met een bloedachtige vloeistof waarin alle organen zijn ingebed; op deze manier kunnen ze gemakkelijk worden voorzien van voedingsstoffen die in het bloed circuleren. Deze vloeistof is kleurloos omdat hij geen pigmenten bevat; om deze reden, het dient slechts een beperkte rol in zuurstoftransport. Twee verschillende typen bloedcellen (of haemocyten) circuleren in de vloeistof: amoebocyten en nefrocyten. De amoebocyten werken waarschijnlijk in bescherming tegen bacteriën en andere vreemde lichamen; bij sommige soorten spelen ze ook een rol in de voortplanting. Nefrocyten absorberen toxines of zetten ze om in een vorm die geschikt is voor eliminatie door de nefridia. De hemocoel wordt door een horizontale verdeling, het middenrif, in twee delen verdeeld: de pericardiale sinus langs de rug en de periviscerale sinus langs de buik. De eerste omsluit het buisvormige hart, en de laatste, de andere organen. Het membraan is op veel plaatsen geperforeerd, waardoor de uitwisseling van vloeistoffen tussen de twee holtes mogelijk is. Het hart zelf is een buis van ringvormige spieren bestaande uit epitheliale weefsels, met twee laterale openingen (ostia) per segment. Hoewel het niet bekend is of de achterkant open of gesloten is, opent deze direct vanaf de voorkant in de lichaamsholte. Aangezien er geen bloedvaten zijn, afgezien van de fijne bloedvaten die lopen tussen de spierlagen van de lichaamswand en een paar slagaders die de antennes voeden, wordt dit aangeduid als een open circulatie. De timing van de pompprocedure kan worden onderverdeeld in twee delen: diastole en systole. Tijdens diastole, bloed stroomt door de ostia van de pericardiale sinus (de holte met het hart) in het hart. Wanneer de systole begint, de Ostia sluiten en de hartspieren samentrekken naar binnen, waardoor het volume van het hart. Dit pompt het bloed van de voorkant van het hart in de periviscerale sinus die de organen bevat. Op deze manier worden de verschillende organen voorzien van voedingsstoffen voordat het bloed uiteindelijk via de perforaties in het middenrif naar de pericardiale sinus terugkeert. Naast de pompende werking van het hart hebben lichaamsbewegingen ook een invloed op de bloedsomloop.
RespirationEdit
zuurstofopname vindt in zekere mate plaats via eenvoudige diffusie door het gehele lichaamsoppervlak, waarbij de coxale blaasjes op de benen mogelijk bij sommige soorten betrokken zijn. Van het grootste belang is echter de uitwisseling van gas via fijne onvertakte buizen, de tracheae, die zuurstof uit het oppervlak tot diep in de verschillende organen, met name het hart, onttrekken. De wanden van deze structuren, die in hun geheel minder dan drie micrometer dik zijn, bestaan slechts uit een extreem dun membraan waardoor zuurstof gemakkelijk kan diffunderen. De tracheae ontstaan bij kleine openingen, de spiracles, die zelf zijn geclusterd in deukachtige uitsparingen van de buitenste huid, de atria. Het aantal aldus gevormde” tracheae-bundels ” bedraagt gemiddeld ongeveer 75 per lichaamssegment; ze hopen zich het dichtst Op Op de rug van het organisme. In tegenstelling tot de geleedpotigen zijn de fluwelen wormen niet in staat om de openingen van hun tracheae te beheersen; de tracheae zijn altijd open, wat in droge omstandigheden een aanzienlijk waterverlies met zich meebrengt. Water gaat twee keer zo snel verloren als bij regenwormen en veertig keer sneller dan bij rupsen. Om deze reden zijn fluweelwormen afhankelijk van habitats met een hoge luchtvochtigheid.
Digestiedit
het spijsverteringskanaal begint iets achter het hoofd, de mond ligt aan de onderzijde een beetje van de voorste punt van het lichaam. Hier kunnen prooien mechanisch door de mandibels met hun bedekking van fijne tandjes worden uiteengereten. Twee speekselklieren lozen via een gemeenschappelijke geleider in de daaropvolgende “keel”, die het eerste deel van de voorste darm vormt. Het speeksel dat zij produceren bevat slijm en hydrolytische enzymen, die de spijsvertering in en buiten de mond in gang zetten. De keel zelf is zeer gespierd en dient om het gedeeltelijk vloeibare voedsel op te nemen en via de slokdarm, die het achterste deel van de voorste darm vormt, naar de centrale darm te pompen. In tegenstelling tot de voorste darm, is dit niet bekleed met een cuticula, maar in plaats daarvan bestaat slechts uit een enkele laag van epitheliaale weefsel, die geen opvallende inkeping vertoont zoals bij andere dieren wordt gevonden. Bij het invoeren van de centrale darm, worden voedseldeeltjes bedekt met een slijm-gebaseerde peritrofische membraan, die dient om de voering van de darm te beschermen tegen schade door scherpe deeltjes. Het intestinale epitheel scheidt verdere spijsverteringsenzymen af en absorbeert de vrijgekomen voedingsstoffen, hoewel de meeste spijsvertering al extern of in de mond heeft plaatsgevonden. Onverteerbare restanten komen in de achterste darm, of rectum, die opnieuw is bekleed met een cuticula en die opent op de anus, gelegen aan de onderzijde in de buurt van de achterkant.
in bijna elk segment is een paar uitscheidingsorganen genaamd nefridia, die zijn afgeleid van coelomweefsel. Elk bestaat uit een klein zakje dat via een gesgelleerde geleider, een nephridioduct genaamd, verbonden is met een opening aan de basis van het dichtstbijzijnde been dat bekend staat als een nephridiopoor. Het zakje wordt ingenomen door speciale cellen genaamd podocyten, die ultrafiltratie van het bloed door de partitie tussen hemocoelom en nefridium vergemakkelijken. De samenstelling van de urinaire oplossing wordt gewijzigd in het nefridioduct door selectieve recuperatie van voedingsstoffen en water en door isolatie van gif en afvalmaterialen, voordat het via de nefridiopoor naar de buitenwereld wordt uitgescheiden. Het belangrijkste stikstofhoudende uitscheidingsproduct is het in water onoplosbare urinezuur; dit kan in vaste toestand met zeer weinig water worden uitgescheiden. Deze zogenaamde uricotelische uitscheidingsmodus vertegenwoordigt een aanpassing aan het leven op het land en de daarmee samenhangende noodzaak om economisch met water om te gaan. Een paar voormalige nefridia in het hoofd werden in de tweede plaats omgezet in de speekselklieren, terwijl een ander paar in het laatste segment van mannelijke specimens nu dienen als klieren die blijkbaar een rol spelen in de voortplanting.
Sensatiedit
het hele lichaam, met inbegrip van de stoppoten, is bezaaid met talrijke papillen: wrattige uitsteeksels reageren op aanraking die een mechanoreceptieve borstel aan het uiteinde dragen, die elk ook is verbonden met verdere sensorische zenuwcellen die eronder liggen. De mondpapillen, de uitgangen van de slijmklieren, hebben waarschijnlijk ook een functie in de zintuiglijke waarneming. Sensorische cellen bekend als” sensills “op de” lippen ” of labrum reageren op chemische stimuli en zijn bekend als chemoreceptoren. Deze zijn ook te vinden op de twee antennes, die de belangrijkste zintuiglijke organen van de fluwelen worm lijken te zijn.
behalve bij enkele (typisch ondergrondse) soorten ligt achter elke antenne een eenvoudig geconstrueerd oog (ocellus), zijdelings, net onder het hoofd. Deze bestaat uit een chitineuze ball lens, een hoornvlies en een netvlies en is verbonden met het centrum van de hersenen via een oogzenuw. Het netvlies bestaat uit talrijke pigmentcellen en fotoreceptoren; deze laatste zijn gemakkelijk aangepaste flagellated cellen, waarvan flagellum membranen een lichtgevoelig pigment op hun oppervlak dragen. De rhabdomere ogen van de Onychophora worden verondersteld homoloog te zijn met de mediane ocelli van geleedpotigen.; dit suggereert dat de laatste gemeenschappelijke voorouder van geleedpotigen alleen mediane ocelli had.De innervatie laat echter zien dat de homologie beperkt is: de ogen van Onychophora vormen zich achter de antenne, terwijl het tegenovergestelde het geval is bij geleedpotigen.
Reproductiedit
beide geslachten bezitten paren van gonaden, die via een kanaal dat een gonoduct wordt genoemd, openen in een gemeenschappelijke genitale opening, de gonopoor, die zich aan de achterste ventrale zijde bevindt. Zowel de gonaden als het gonoduct zijn afgeleid van echt coelomweefsel.
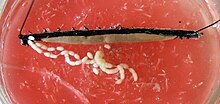
bij vrouwtjes zijn de twee eierstokken in het Midden en aan het horizontale middenrif verbonden. Het gonoduct lijkt anders, afhankelijk van of de soort levend-dragend of ei-leggend is. Bij levend dragende soorten verdeelt elk uitgangskanaal zich in een slanke ovidict en een ruime “baarmoeder”, de baarmoeder, waarin de embryo ‘ s zich ontwikkelen. De enkele vagina, waaraan beide baarmoeder zijn verbonden, loopt naar buiten naar de gonopoor. Bij eierleggende soorten, waarvan het gonoduct uniform is geconstrueerd, ligt de genitale opening aan de punt van een lang eilegapparaat, de legboor. De vrouwtjes van veel soorten bezitten ook een spermacentrum, de opvangbak seminis, waarin spermacellen van mannetjes tijdelijk of langer bewaard kunnen worden.
mannetjes bezitten twee afzonderlijke testes, samen met de bijbehorende zaadblaasjes (de vesicula seminalis) en het uitgangskanaal (de vasa efferentia). De twee Vasa efferentia verenigen zich in een gemeenschappelijke zaadbuis, de vas deferens, die op zijn beurt verbreedt door het ejaculatory kanaal te openen bij de gonopoor. Direct naast of achter deze liggen twee paar speciale klieren, die waarschijnlijk een extra voortplantingsfunctie hebben; de achterste klieren zijn ook bekend als anale klieren. Een penis-achtige structuur is tot nu toe alleen gevonden bij mannetjes van het geslacht Paraperipatus, maar is nog niet waargenomen in actie.
Er zijn verschillende paringsprocedures: Bij sommige soorten plaatsen mannetjes hun spermatofoor direct in de genitaliën van het wijfje, terwijl anderen het op het wijfje lichaam deponeren, waar de cuticula zal instorten en waardoor de zaadcellen in het wijfje kunnen migreren. Er zijn ook Australische soorten waar het mannetje zijn spermatofoor op zijn kop plaatst, die dan tegen de geslachtsdelen van het vrouwtje wordt gedrukt. Bij deze soorten heeft het hoofd uitgebreide structuren zoals spikes, stekels, holle stylets, putten, en depressies, waarvan het doel is om ofwel het sperma te houden en / of te helpen bij de sperma-overdracht aan het vrouwtje. De mannetjes van de meeste soorten scheiden ook een feromoon af van klieren aan de onderzijde van de poten om vrouwtjes aan te trekken.