(„człowiek” / „gwiazda” w języku Sotho)
strona
Rising Star cave system, RPA
ludzie
Jaskiniowcy Steven Tucker i Rick Hunter oraz badani przez Paula Dirksa i Lee Bergera i ich współpracowników

wprowadzenie
ten najnowszy członek naszego rodzaju po raz kolejny pomieszał ewolucyjną historię linii Homo. Najbardziej ekscytującym aspektem jest natura szczątków sugeruje, że zostały celowo złożone w głębokiej pieczarze, gdzie zostały odkryte. H. heidelbergensis był do tej pory najwcześniejszym gatunkiem uważanym za uprawiający celowe usuwanie ciała. Próby datowania szczątków nie powiodły się. Thackeray (2015) oszacował jednak, że gatunek może datować się na 2.0 ± 0,5 mya, na podstawie porównań daty i cech anatomicznych wśród H. naledi, H. habilis, H. rudolfensis i H. erectus (patrz rysunek 26.2 i Rozdział 27 dla klasy erectus).
filogeneza
wydaje się, że większość badaczy zgadza się, że szczątki odzwierciedlają nowy hominin (patrz odniesienia w tej sekcji, zwłaszcza Randolph-Quinney 2015). Podobnie jak większość homininów, filogeneza gatunku jest nieznana, ale prawdopodobnie pochodzi od australopitów. Co jeszcze bardziej utrudnia to fakt, że gatunek ten dzieli cechy z możliwymi istniejącymi lub prawie istniejącymi gatunkami Homo (H. habilis, H. rudolfensis i H. erectus), bardziej pochodnymi formami (np. neandertalczycy i ludzie), a także różnymi australopitami. Mozaika cech jest interesująca i dodatkowo wspiera krzaczasty charakter drzewa homininowego.
odkrycie i zasięg geograficzny
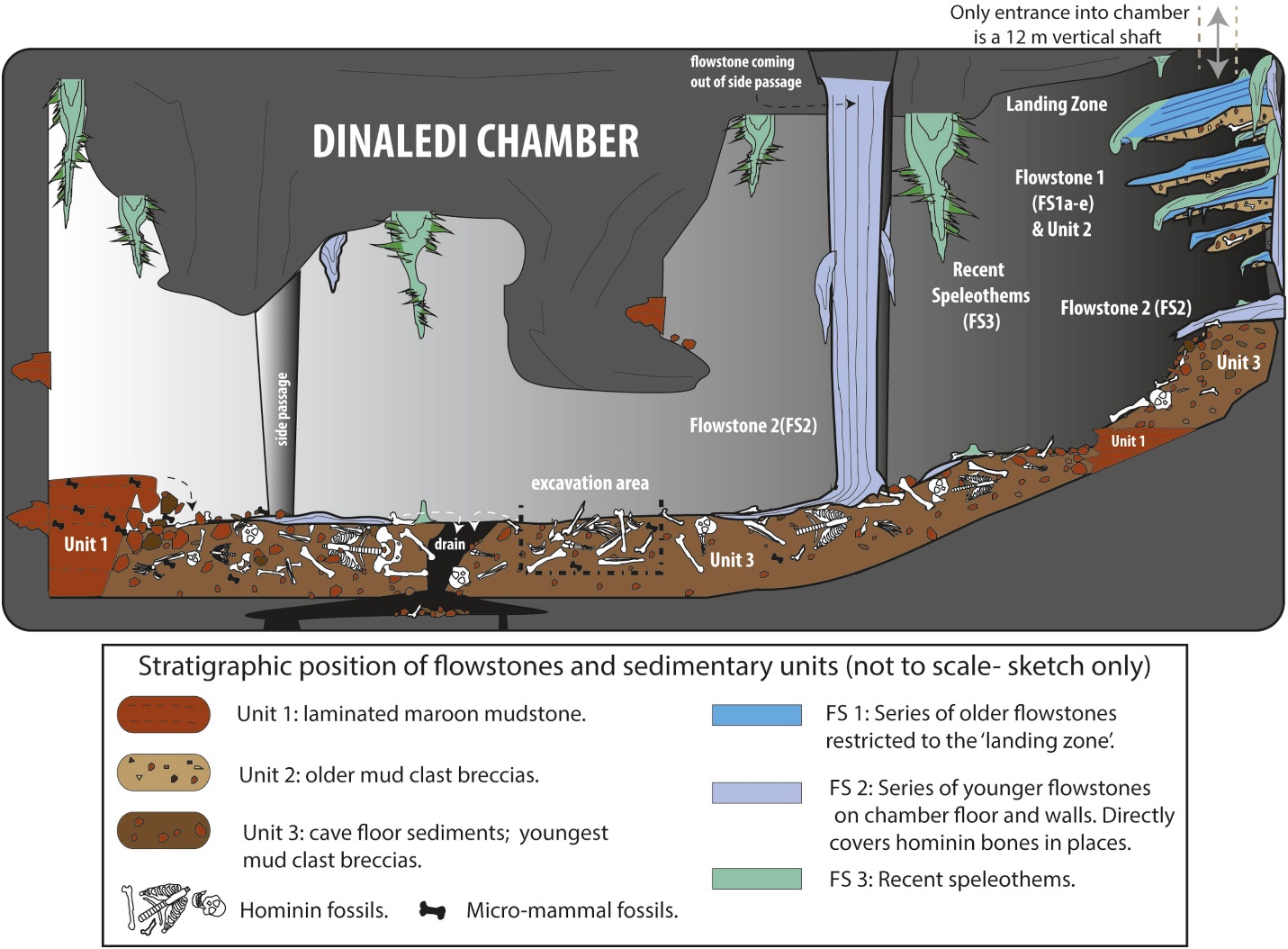
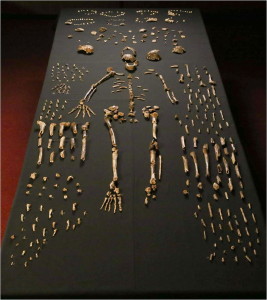
szczątki minimum 15 osobników, łącznie 1550 skamieniałości (patrz rysunek 26.4), zostały wydobyte w 2013 i 2014 roku z komory Dinaledi, znajdującej się w systemie jaskiń wschodzącej gwiazdy w Cradle of Humankind World Heritage Site, Prowincja Gauteng, RPA (Berger et al. 2015). Skamieniałości są największym zbiorem gatunków hominidów w Afryce (Dirks et al. 2015). Komora znajduje się 30 m pod ziemią i jest dostępna tylko przez wąski szyb o długości 12 m (Randolph-Quinney 2015 i patrz rysunek 26.3 – prawy górny róg). Na podstawie danych depozytowych ciała zostały zdeponowane w czasie (Dirks et al. 2015).
cechy fizyczne
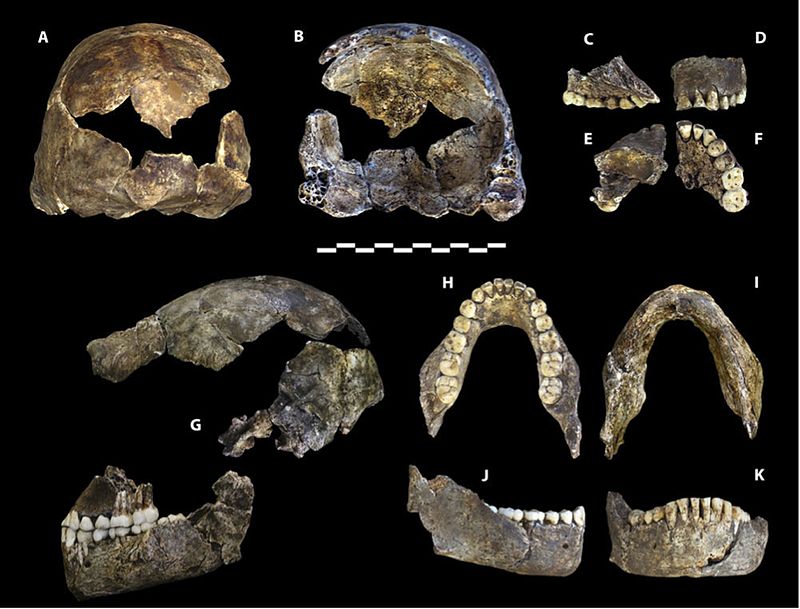
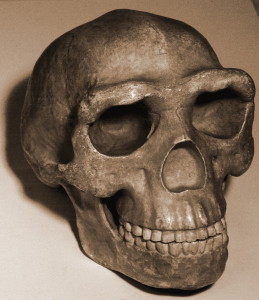
szczątki są szczególnie cenne, ponieważ reprezentowane są wszystkie regiony ciała, a niektóre kości są przegubowe, dzięki czemu zachowane są anatomiczne pozycje i układy, np. prawie kompletna noga dziecka i Dorosła ręka (Dirks et al. 2015). Niska pojemność czaszki, podwyższone stawy barkowe, zakrzywione paliczki oraz morfologia tułowia i biodra są podobne do australopitów. Morfologia czaszki, szczęki i zębów oraz kości nóg są w większości Homopodobne. Nadgarstek jest najbardziej podobny do ludzi i neandertalczyków. Stopa jest bardzo ludzka. (Berger et al. 2015, Harcourt-Smith et al. 2015, Kivell 2015, Thackeray 2015) w ten sposób widzimy zdolnego dwunożnego lądowego, który może wspinać się, żerować i schronić się na drzewach.
pojemność czaszki mieści się w zakresie dla australopitów, przy czym samce osiągają średnio 560 cm3, a samice 465 cm3 (Berger et al. 2015). Podstawa sklepienia czaszki jest wygięta jak członkowie klasy erectus i kolejnych gatunków Homo (patrz rysunek 26.2 i 15.8). Kości sklepienia są cienkie jak u H. habilis. H. naledi wykazuje mniejsze zwężenie postorbitalne niż najwcześniejsze australopity, ale posiada większy torus nadoczodołowy niż jakikolwiek australopit gracile(Berger et al. 2015). Razem wzięte, jest to dziwna kombinacja. Bardziej zgrabna twarz spowodowałaby mniejsze zwężenie postorbitalne, ale torus nadoczodołowy jest związany ze stresem żucia. Co bardziej zaskakujące, zęby są mniejsze niż zęby H. habilis, H. rudolfensis i klasy erectus, z wyjątkiem H. floresiensis(Berger et al. 2015). Sklepienie czaszki jest pięciokątne w przekroju jak Azjatycki H. erectus (patrz rysunek 26.5), ze względu na obecność stępki strzałkowej, tj. zgrubienia wzdłuż linii środkowej czaszki, od przodu do tyłu (Berger et al. 2015). Ponieważ cecha jest słabo poznana w H. erectus, jego obecność u innego gatunku może rzucić więcej światła na znaczenie adaptacyjne lub procesy ontologiczne. Jednak, w przeciwieństwie do H. erectus, kil zaczyna się w ciemieniowej w porównaniu do obszaru czołowego (ponownie, patrz rysunek 26.5). Chociaż nie jest dobrze rozwinięty, H. naledi wykazuje fossa psa, czyli depresja powyżej zęba psa, jak widać u H. antecessor, H. heidelbergensis i ludzi.
podczas gdy ręka H. naledi (patrz rysunek 26.6) ma cechy wspólne z innymi homininami, połączenie cech jest unikalne. Miały długie palce, a dwa bliższe paliczki palców są zakrzywione nawet bardziej niż u australopitów, co sugeruje aktywność nadrzewną. Jednak ich morfologia nadgarstka jest najbardziej podobna do neandertalczyków i współczesnych ludzi, a wraz z ich długim, solidnym kciukiem byli w ten sposób zdolni do silnych działań manipulacyjnych (Berger et al. 2015, Kivell et al. 2015).
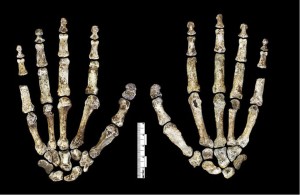
Klatka piersiowa i miednica były rozszerzone jak australopity, ale kręgi przypominały kręgi klasy erectus i kolejnych gatunków Homo.
podczas gdy połączenie cech występujących w kościach nóg jest charakterystyczne, są one Homo-podobne, z tym, że szyja udowa jest długa jak u australopitów. Stopa (patrz rysunek 26.7) jest bardzo podobny do człowieka, z podstawowymi różnicami są krzywizna ich cyfr i mniej przyśrodkowego łuku podłużnego (Harcourt-Smith 2015).
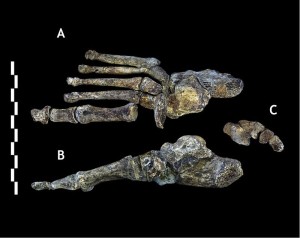
pozornie zaawansowana dwunożna morfologia dwóch ostatnich odkryć hominin, tj. Au. sediba i H. naledi sugerują, że silne siły selektywne sprzyjały zdolności poruszania się po krajobrazie, w poszukiwaniu żywności i innych zasobów.
(2015) obliczyli wagę płci w następujący sposób: samce na 55,8 kg i samice na 39,7 kg. Jedyną kość, której mogli użyć do obliczenia wysokości, była kość piszczelowa, która dała oszacowanie 144,5-147,8 mm. Stwierdzają, że gatunek mieści się w zakresie wysokości współczesnych małych populacji ludzkich, jak również szacunki dla H. georgicus hypodigm z Dmanisi(Lordkipanidze et al. 2007, cytowany w Berger et al. 2015).
środowisko i sposób życia
środowisko regionu, około 2 mln lat temu, zostało już opisane w sekcji „środowisko i sposób życia” Au. sediba i P. robustus. Co ciekawe, stanowisko Dinaledi znajduje się 800 m na południowy zachód od stanowiska P. robustus w Swartkrans. To była dzielnica hominin!
dowody są przekonujące, że szczątki nie mogły zostać złożone za pomocą sił naturalnych, ale raczej zostały przeniesione przynajmniej w części drogi, przez ciemne i wąskie przejście. Musimy więc ponownie ocenić nasz obraz zdolności poznawczych i świadomości wcześniejszych członków naszego rodzaju.