(« humain”/ »étoile” en langue Sotho)
SITE
Rising Star cave system, Afrique du Sud
PERSONNES
Spéléologues Steven Tucker et Rick Hunter et étudiés par Paul Dirks et Lee Berger et leurs associés
INTRODUCTION
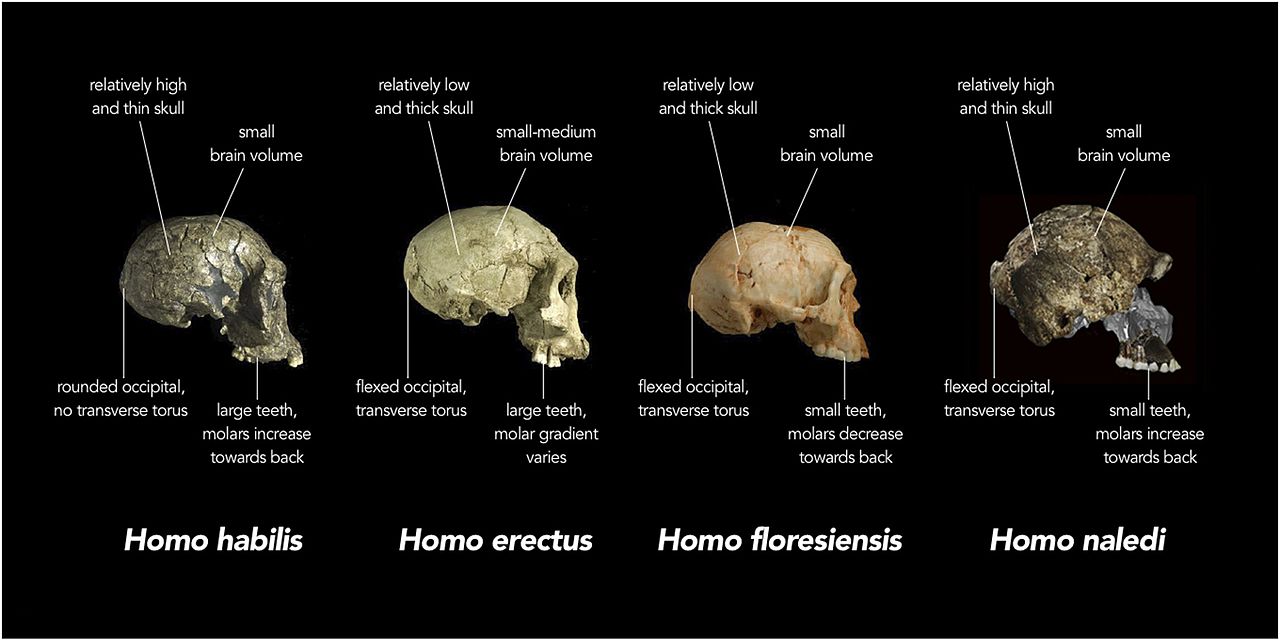
Ce nouveau membre de notre genre a une fois de plus confondu l’histoire évolutive de la lignée Homo. L’aspect le plus excitant est que la nature des restes suggère qu’ils ont été déposés intentionnellement dans la caverne profonde où ils ont été découverts. H. heidelbergensis était jusqu’ici la première espèce à avoir pratiqué l’élimination intentionnelle du corps. Les tentatives de datation des restes n’ont pas abouti. Cependant, Thackeray (2015) a estimé que l’espèce pourrait dater de 2.0 ± 0,5 mya, basé sur des comparaisons de dates et de caractéristiques anatomiques entre H. naledi, H. habilis, H. rudolfensis et H. erectus (voir la figure 26.2 et le chapitre 27 pour le grade erectus).
PHYLOGÉNIE
Il semble que la majorité des chercheurs s’accordent à dire que les restes reflètent un nouvel hominine (voir les références à cette section, en particulier Randolph-Quinney 2015). Comme la plupart des hominins, la phylogénie de l’espèce est inconnue, mais elle descend probablement d’une ascendance australopithèque. Ce qui rend les choses encore plus difficiles, c’est que l’espèce partage des caractéristiques avec d’éventuelles espèces existantes ou proches d’Homo (H. habilis, H. rudolfensis et H. erectus), des formes plus dérivées (par exemple, les néandertaliens et les humains), ainsi que divers australopithes. La mosaïque de traits est intéressante et soutient davantage la nature touffue de l’arbre hominin.
DÉCOUVERTE ET RÉPARTITION GÉOGRAPHIQUE
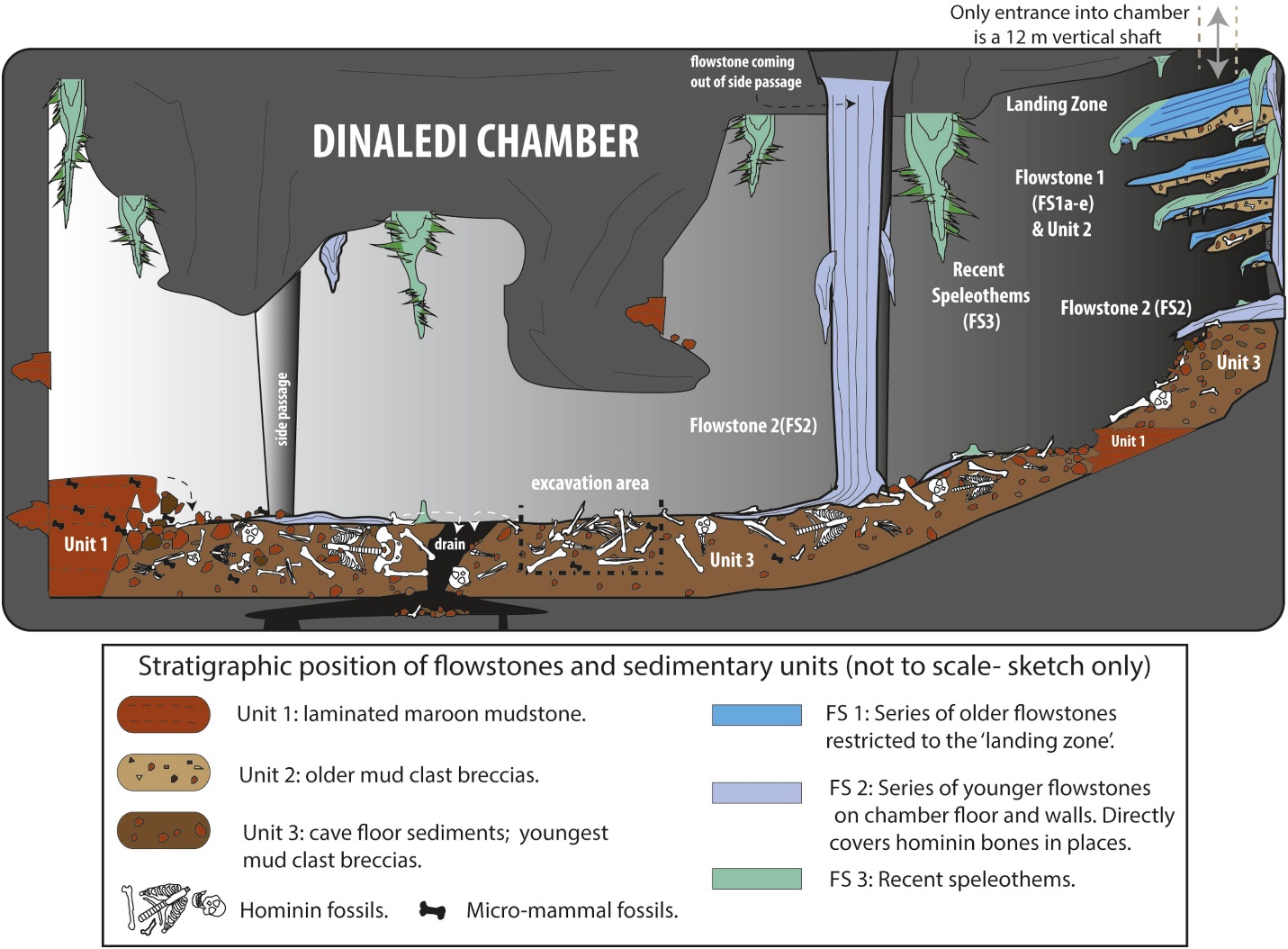
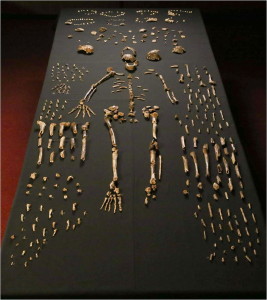
Les restes d’un minimum de 15 individus, totalisant 1550 fossiles (voir Figure 26.4), ont été fouillées en 2013 et 2014 dans la Chambre de Dinaledi, située dans le système de grottes de l’Étoile Montante dans le Site du Patrimoine mondial du Berceau de l’Humanité, dans la province de Gauteng, en Afrique du Sud (Berger et al. 2015). Les fossiles constituent la plus grande collection d’une espèce d’hominidés en Afrique (Dirks et al. 2015). La chambre se trouve à 30 m sous terre et n’est accessible que par un puits étroit de 12 m (Randolph-Quinney 2015 et voir la figure 26.3 – en haut à droite). Sur la base des données de dépôt, les corps ont été déposés au fil du temps (Dirks et al. 2015).
CARACTÉRISTIQUES PHYSIQUES
Les restes sont particulièrement précieux car toutes les régions du corps sont représentées et certains os sont articulés, de sorte que les positions et les dispositions anatomiques sont préservées, par exemple une jambe presque complète d’un enfant et une main adulte (Dirks et al. 2015). La faible capacité crânienne, les articulations de l’épaule surélevées, les phalanges incurvées et la morphologie du tronc et de la hanche ressemblent à des australopithèques. La morphologie de la crane, de la mâchoire et des dents, ainsi que les os des jambes sont, pour la plupart, de type Homo. Le poignet ressemble le plus aux humains et aux néandertaliens. Le pied est très humain. (Berger et coll. 2015, Harcourt-Smith et coll. 2015, Kivell 2015, Thackeray 2015) Ainsi, nous voyons un bipède terrestre capable de grimper, de se nourrir et de se réfugier dans les arbres.
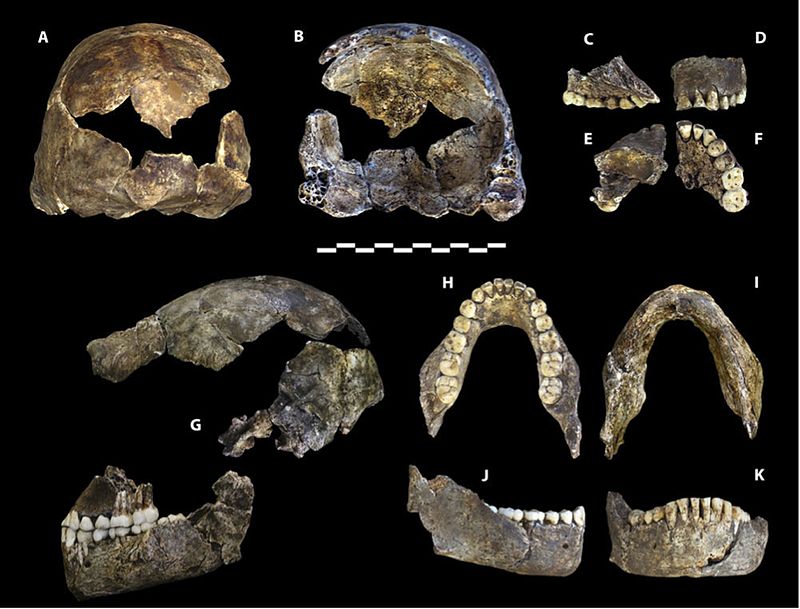
La capacité crânienne se situe dans la plage des australopithèques, les mâles mesurant en moyenne 560 cm3 et les femelles 465 cm3 (Berger et al. 2015). La base de la voûte crânienne est fléchie comme les membres du grade erectus et les espèces suivantes d’Homo (voir Figures 26.2 et 15.8). Les os de la voûte sont minces comme ceux de H. habilis. H. naledi présente moins de constriction postorbitale que les premiers australopithes, mais possède un tore supraorbital plus grand que n’importe quel australopithe gracile (Berger et al. 2015). Pris ensemble, c’est une combinaison étrange. Un visage plus gracile entraînerait moins de constriction postorbitale, mais le tore supraorbital est associé à un stress de mastication. Plus surprenant encore, les dents sont plus petites que celles de H. habilis, H. rudolfensis et du grade erectus, à l’exception de H. floresiensis (Berger et al. 2015). La voûte crânienne est de section pentagonale comme le H. erectus asiatique (voir Figure 26.5), en raison de la présence d’une quille sagittale, c’est-à-dire d’un épaississement le long de la ligne médiane du crâne, de l’avant vers l’arrière (Berger et al. 2015). Puisque le trait est mal compris dans H. erectus, sa présence chez une autre espèce peut éclairer davantage la signification adaptative ou les processus ontologiques impliqués. Cependant, contrairement à H. erectus, la quille commence dans la région pariétale par rapport à la région frontale (encore une fois, voir Figure 26.5). Bien que peu développé, H. naledi présente une fosse canine, c’est-à-dire une dépression au-dessus de la dent canine, comme on le voit chez H. antecessor, H. heidelbergensis et chez l’homme.
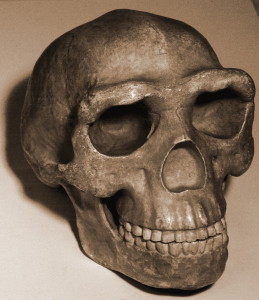
Bien que la main de H. naledi (voir Figure 26.6) partage des caractéristiques avec d’autres hominines, la combinaison de caractéristiques est unique. Ils avaient de longs doigts et les deux phalanges plus proximales sont courbées encore plus que celles des australopithes, suggérant des activités arboricoles. Pourtant, leur morphologie du poignet est très similaire à celle des néandertaliens et des humains modernes et, avec leur pouce long et robuste, ils étaient donc capables de fortes activités de manipulation (Berger et al. 2015, Kivell et coll. 2015).
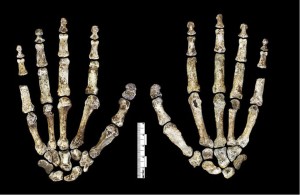
Le thorax et le bassin étaient évasés comme des australopithes, mais les vertèbres ressemblent à celles du grade erectus et des espèces d’Homo suivantes.
Bien que la combinaison des caractéristiques observées dans les os de la jambe soit distinctive, elles ressemblent à des Homo, sauf que le col fémoral est long comme celui des australopithes. Le pied (voir Figure 26.7) est très semblable à l’homme, les principales différences étant la courbure de leurs doigts et moins une arche longitudinale médiale (Harcourt-Smith 2015).
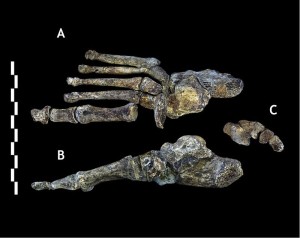
La morphologie bipède apparemment avancée des deux découvertes les plus récentes d’hominines, c’est-à-dire Au. sediba et H. naledi suggèrent que de fortes forces sélectives favorisaient la capacité de se déplacer dans le paysage, à la recherche de nourriture et d’autres ressources.
Berger et coll. (2015) ont calculé le poids des sexes comme suit: les mâles à 55,8 kg et les femelles à 39,7 kg. Le seul os qu’ils pouvaient utiliser pour calculer la hauteur était un tibia qui donnait une estimation de 144,5 à 147,8 mm. Ils indiquent que l’espèce se situe dans la plage de hauteur des populations humaines modernes à petite taille, ainsi que des estimations pour l’hypodigme H. georgicus de Dmanisi (Lordkipanidze et al. 2007, cité dans Berger et al. 2015).
ENVIRONNEMENT ET MODE DE VIE
L’environnement de la région, à environ 2 mia, a déjà été décrit dans les sections « ENVIRONNEMENT ET MODE DE VIE » de l’Ua. sediba et P. robustus. Fait intéressant, le site de Dinaledi se trouve à 800 m au sud-ouest du site de P. robustus de Swartkrans. C’était un quartier d’hominins !
La preuve est convaincante que les restes n’auraient pas pu être déposés par des forces naturelles, mais ont plutôt été transportés au moins en partie, à travers un passage sombre et étroit. Nous devons donc réévaluer notre image des capacités cognitives et de la conscience des membres antérieurs de notre genre.