(„Mensch“ / „Stern“ in Sotho-Sprache)
WEBSITE
Rising Star Cave system, Südafrika
MENSCHEN
Höhlenforscher Steven Tucker und Rick Hunter und untersucht von Paul Dirks und Lee Berger und ihre Mitarbeiter
EINFÜHRUNG
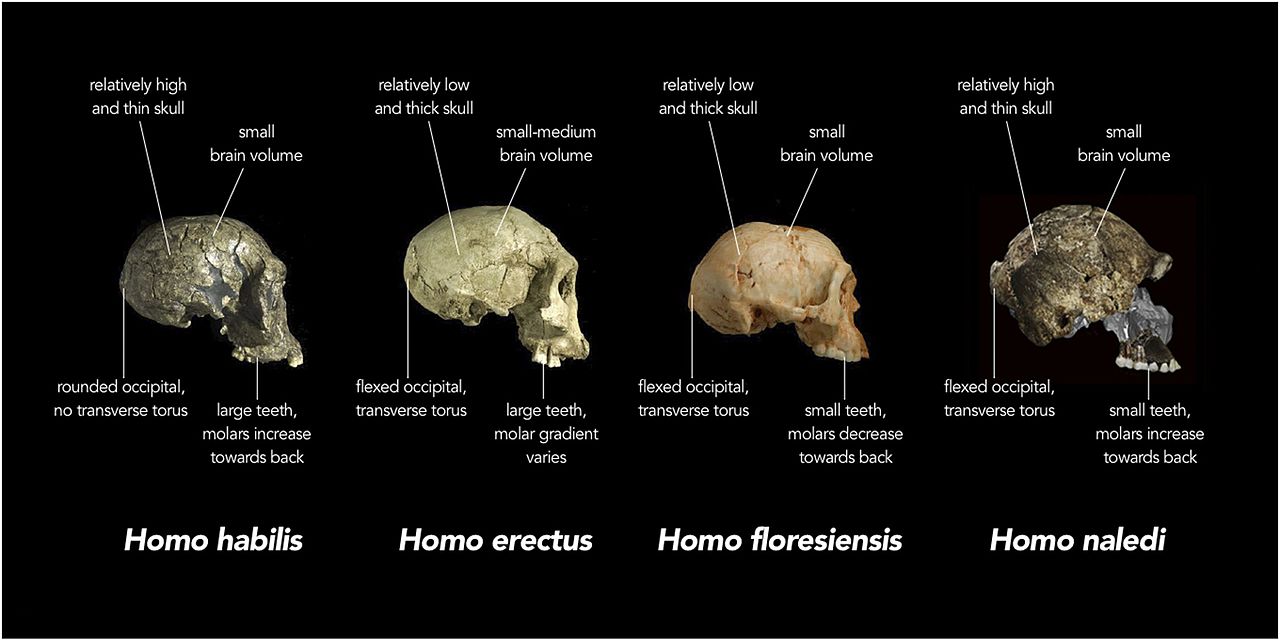
Dieses neueste Mitglied unserer Gattung hat die Evolutionsgeschichte der Homo-Linie erneut durcheinander gebracht. Der aufregendste Aspekt ist, dass die Art der Überreste darauf hindeutet, dass sie absichtlich in der tiefen Höhle abgelagert wurden, in der sie entdeckt wurden. H. heidelbergensis war bisher die früheste Spezies, von der angenommen wurde, dass sie absichtliche Körperentsorgung praktiziert hat. Versuche Dating Die Überreste waren nicht erfolgreich. Thackeray (2015) hat jedoch geschätzt, dass die Art auf 2 datieren kann.0 ± 0,5 mya, basierend auf Vergleichen von Datum und anatomischen Merkmalen zwischen H. naledi, H. habilis, H. rudolfensis und H. erectus (siehe Abbildung 26.2 und Kapitel 27 für den erectus-Grad).
PHYLOGENIE
Es scheint, dass die Mehrheit der Forscher zustimmt, dass die Überreste ein neues Hominin widerspiegeln (siehe Referenzen in diesem Abschnitt, insbesondere Randolph-Quinney 2015). Wie die meisten Homininen ist die Phylogenie der Art unbekannt, aber sie stammt wahrscheinlich von einer australopithischen Abstammung ab. Was die Dinge noch schwieriger macht, ist, dass die Art Merkmale mit möglichen vorhandenen oder nahezu vorhandenen Homo-Arten (H. habilis, H. rudolfensis und H. erectus), abgeleiteten Formen (z. B. Neandertaler und Menschen) sowie verschiedenen Australopithen teilt. Das Mosaik der Merkmale ist interessant und unterstützt die buschige Natur des Homininbaums.
ENTDECKUNG UND GEOGRAFISCHE REICHWEITE
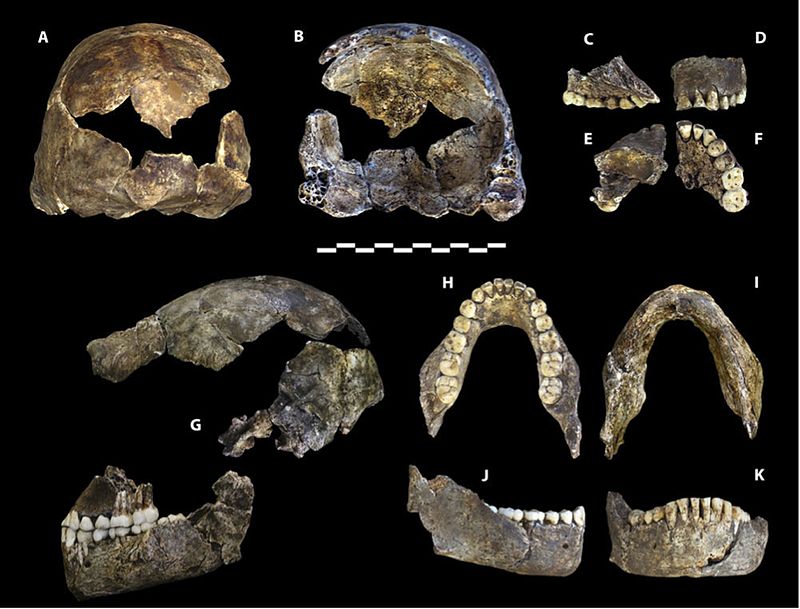
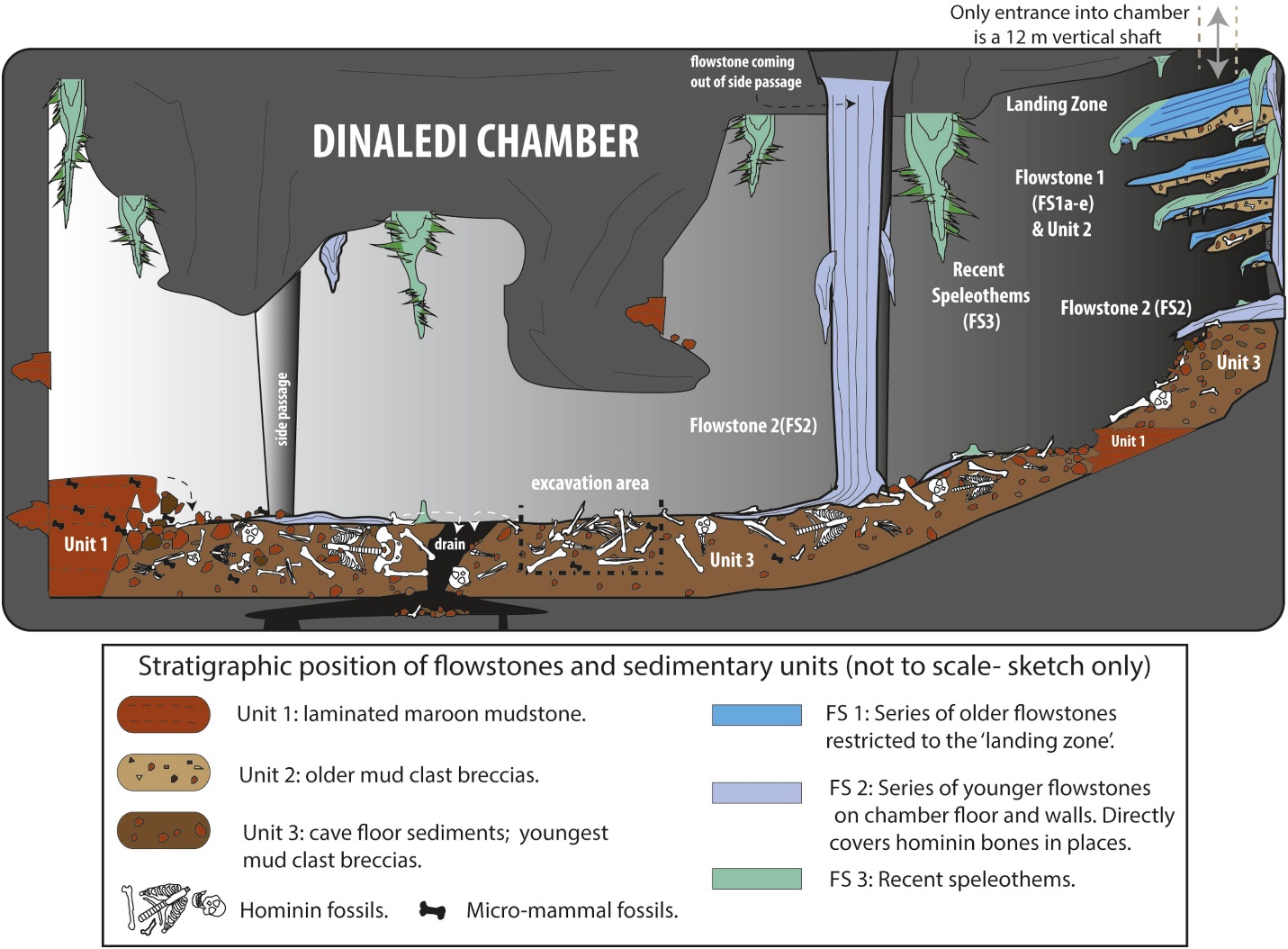
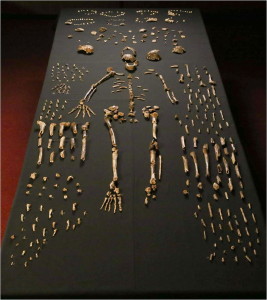
Die Überreste von mindestens 15 Individuen, insgesamt 1550 Fossilien (siehe Abbildung 26.4), wurden 2013 und 2014 aus der Dinaledi-Kammer ausgegraben, die sich im Rising Star Cave System im Weltkulturerbe Cradle of Humankind in der Provinz Gauteng, Südafrika, befindet (Berger et al. 2015). Die Fossilien sind die größte Sammlung einer Hominidenart in Afrika (Dirks et al. 2015). Die Kammer liegt 30 m unter der Erde und ist nur über einen 12 m schmalen Schacht zugänglich (Randolph-Quinney 2015 und siehe Abbildung 26.3 – oben rechts). Basierend auf Ablagerungsdaten wurden die Körper im Laufe der Zeit abgelagert (Dirks et al. 2015).
PHYSIKALISCHE EIGENSCHAFTEN
Die Überreste sind besonders wertvoll, da alle Körperregionen dargestellt sind und einige Knochen artikuliert sind, so dass anatomische Positionen und Anordnungen erhalten bleiben, z. B. ein fast vollständiges Bein eines Kindes und eine erwachsene Hand (Dirks et al. 2015). Die niedrige Schädelkapazität, die erhöhten Schultergelenke, die gekrümmten Phalangen sowie die Rumpf- und Hüftmorphologie sind australopithenähnlich. Crania, Kiefer- und Zahnmorphologie, und Beinknochen sind, hauptsächlich, Homo-ähnlich. Das Handgelenk ist Menschen und Neandertalern am ähnlichsten. Der Fuß ist sehr menschenähnlich. (Berger et al. 2015, Harcourt-Smith et al. 2015, Kivell 2015, Thackeray 2015) So sehen wir einen fähigen terrestrischen Zweibeiner, der klettern, Futter suchen und in Bäumen Zuflucht suchen kann.
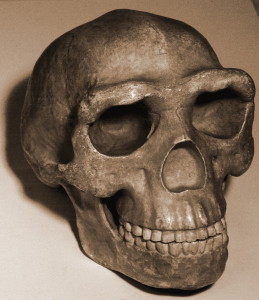
Die Schädelkapazität liegt im Bereich der Australopithe, wobei die Männchen durchschnittlich 560 ccm und die Weibchen 465 ccm haben (Berger et al. 2015). Die Basis des Schädelgewölbes ist wie bei Mitgliedern der Erectus-Klasse und nachfolgenden Homo-Arten gebogen (siehe Abbildung 26.2 und 15.8). Die Gewölbeknochen sind dünn wie die von H. habilis. H. naledi weist eine geringere Postorbitalverengung auf als die frühesten Australopithe, besitzt jedoch einen größeren supraorbitalen Torus als jeder grazile Australopith (Berger et al. 2015). Zusammengenommen ist es eine seltsame Kombination. Ein grazileres Gesicht würde zu einer geringeren Postorbitalverengung führen, der supraorbitale Torus ist jedoch mit Kaustress verbunden. Noch überraschender ist, dass die Zähne kleiner sind als die von H. habilis, H. rudolfensis und der erectus-Klasse, mit Ausnahme von H. floresiensis (Berger et al. 2015). Das Schädelgewölbe ist im Querschnitt fünfeckig wie der asiatische H. erectus (siehe Abbildung 26.5), da ein sagittaler Kiel vorhanden ist, d. H. Eine Verdickung entlang der Mittellinie des Schädels von vorne nach hinten (Berger et al. 2015). Da das Merkmal in H schlecht verstanden wird. erectus, seine Anwesenheit in einer anderen Spezies kann mehr Licht auf die adaptive Bedeutung oder ontologische Prozesse werfen. Im Gegensatz zu H. erectus beginnt der Kiel jedoch im parietalen Bereich gegenüber dem frontalen Bereich (siehe Abbildung 26.5). H. naledi ist zwar nicht gut entwickelt, weist jedoch eine Fossa canina auf, d. H. Eine Vertiefung über dem Eckzahn, wie bei H. antecessor, H. heidelbergensis und Menschen.
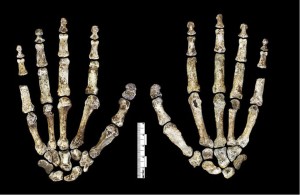
Während die Hand von H. naledi (siehe Abbildung 26.6) Eigenschaften mit anderen Homininen teilt, ist die Kombination von Eigenschaften einzigartig. Sie hatten lange Finger und die beiden proximalen Fingerphalangen sind noch stärker gekrümmt als die von Australopiths, was auf Baumaktivitäten hindeutet. Ihre Handgelenkmorphologie ist jedoch den Neandertalern und modernen Menschen am ähnlichsten und sie waren zusammen mit ihrem langen, robusten Daumen zu starken Manipulationsaktivitäten fähig (Berger et al. 2015, Kivell et al. 2015).
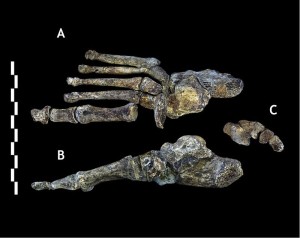
Der Thorax und das Becken waren wie Australopithe ausgestellt, aber die Wirbel ähneln denen der Erectus-Klasse und nachfolgender Homo-Arten.
Während die Kombination von Merkmalen, die in den Beinknochen zu sehen sind, unverwechselbar ist, sind sie Homo-ähnlich, außer dass der Schenkelhals lang ist wie der von Australopiths. 26.7) ist sehr menschenähnlich, wobei die Hauptunterschiede in der Krümmung ihrer Ziffern und weniger in einem medialen Längsgewölbe bestehen (Harcourt-Smith 2015).
Die scheinbar fortgeschrittene bipedale Morphologie der beiden jüngsten Hominin-Entdeckungen, d.h. Au. sediba und H. naledi schlagen vor, dass starke selektive Kräfte die Fähigkeit begünstigten, sich auf der Suche nach Nahrung und anderen Ressourcen durch die Landschaft zu bewegen.
Berger et al. (2015) haben das Gewicht der Geschlechter wie folgt berechnet: Männchen bei 55,8 kg und Weibchen bei 39,7 kg. Der einzige Knochen, den sie zur Berechnung der Höhe verwenden konnten, war eine Tibia, die eine Schätzung von 144,5 – 147,8 mm ergab. Sie geben an, dass die Art in den Höhenbereich moderner kleiner menschlicher Populationen fällt, sowie Schätzungen für den H. georgicus hypodigm aus Dmanisi (Lordkipanidze et al. 2007, zitiert in Berger et al. 2015).
UMWELT UND LEBENSWEISE
Die Umwelt der Region, bei etwa 2 mya, wurde bereits in den Abschnitten „UMWELT UND LEBENSWEISE“ von Au beschrieben. sediba und P. robustus. Interessanterweise liegt das Dinaledi-Gelände 800 m südwestlich des P. robustus-Geländes von Swartkrans. Es war eine hominin Nachbarschaft!Die Beweise sind überzeugend, dass die Überreste nicht durch natürliche Kräfte abgelagert worden sein könnten, sondern zumindest einen Teil des Weges durch einen dunklen und engen Durchgang getragen wurden. Wir müssen daher unser Bild von den kognitiven Fähigkeiten und dem Bewusstsein früherer Mitglieder unserer Gattung neu bewerten.