twee van de meest opwindende en veelzijdige genetische hulpmiddelen die de laatste 30 jaar zijn ontwikkeld, zijn de CRE-lox-en FLP-FRT-technologieën. Beide staan de plaats en de timing van genuitdrukking toe om dicht worden geregeld. Dit artikel schetst in het kort hoe de twee systemen werken en kunnen worden gebruikt in muismodellen.de Cre-lox-technologie werd in de jaren tachtig geïntroduceerd (Sauer en Henderson 1988; Sternberg en Hamilton 1981) en werd gepatenteerd door DuPont Pharmaceuticals. Het is met succes toegepast in gisten, planten, zoogdiercelculturen en muizen (Araki et al. 1987). Het is gebaseerd op de capaciteit van de nieuwe combinatie (Cre) recombinase gen (cre) van de bacteriofaagcyclisatie om nieuwe combinatie tussen paren van loxp plaatsen tot stand te brengen. Dergelijke recombinatie in een” Cre-lox ” muis (zie hieronder) kan een gen van belang activeren of inactiveren.
om Cre-lox-technologie te gebruiken, moet een onderzoeker een cre-lox-muis produceren, meestal door een Cre-muis te fokken naar een loxP-muis. Een cre-muis bevat een cre recombinase transgene onder de leiding van een weefsel-specifieke promotor; een loxP-muis bevat twee loxp-plaatsen die een genomisch segment van belang flankeren, de “floxed” locus. Typisch, Cre en loxP muizen worden geproduceerd door middel van transgene technologie (nagy 2000). Afhankelijk van de promotors en andere regulerende controles die worden gebruikt om ze te construeren, kunnen Cre-muizen alleen worden ontworpen om cre-recombinase tot expressie te brengen onder bepaalde voorwaarden, waaronder de volgende:: in bepaalde weefsels, wanneer het dieet van een muis wordt aangevuld met stoffen zoals doxycycline, tetracycline, RU486 en tamoxifen (Brocard et al. 1998; Kellendonk et al. 1999; Utomo et al. 1999) en tijdens bepaalde ontwikkelingsstadia.
afhankelijk van de locatie en oriëntatie van de loxp-locaties in een Cre-lox-muis, kan cre recombinase deleties, inversies en translocaties van de floxed locus initiëren (Nagy 2000) (Fig. 1).
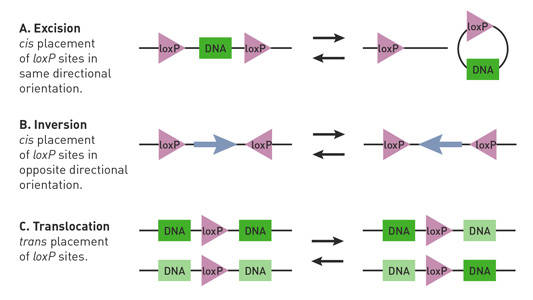 Fig. 1.
Fig. 1.
Cre-lox reacties worden beïnvloed door de oriëntatie en locatie van loxP plaatsen. De in paren gerangschikte loxP plaatsen (driehoeken) hebben directionaliteit en kunnen in een cis (zelfde bundel van DNA) of trans (verschillende bundels van DNA) regeling worden geplaatst. (A) als de loxp-plaatsen een DNA-segment (rechthoek) in een cis-regeling flankeren en in dezelfde richting worden georiënteerd, bemiddelt cre recombinase excisie of circularisatie van het segment. (B) als de loxp-plaatsen het segment van DNA in een cis-regeling flankeren en in tegenovergestelde richtingen worden georiënteerd, bemiddelt cre recombinase de inversie van het segment. (C) als de loxp-plaatsen zich op verschillende bundels van DNA bevinden en in dezelfde richting worden georiënteerd, bemiddelt cre recombinase een translocatie van het segment.
gewoonlijk worden Cre-en loxP-stammen onafhankelijk ontwikkeld en vervolgens gekruist. Veel verschillende Cre-stammen, die elk een Cre transgene bevatten onder leiding van een andere weefselspecifieke promotor, kunnen worden gekruist met één enkele loxp-stam. Afhankelijk van welke stammen worden gedekt, kan een verscheidenheid van Cre-gemedieerde modelsystemen worden geconstrueerd, met inbegrip van transgenics, knockouts, hypomorfen, herstelbare hypomorfen, chromosoomafwijkingen en dieet-geïnduceerde mutanten. In feite, door het mengen en matchen van CRE en loxP stammen, kan een onderzoeker de effecten van een gen bestuderen in weefsel-specifieke en ontwikkelingsstadium-specifieke manieren die voorheen onmogelijk waren.
het FLP-FRT-systeem
het FLP-FRT-systeem is vergelijkbaar met het Cre-lox-systeem en wordt steeds vaker gebruikt in op muizen gebaseerd onderzoek. Het impliceert gebruikend flippase (FLP) recombinase, uit de gist Saccharomyces cerevisiae wordt afgeleid (Sadowski 1995). FLP erkent een paar van de opeenvolgingen van het het doel (FRT) van FLP recombinase die een genomic gebied van belang flankeren.
Araki K, Imaizumi T, Okuyama K, Oike Y, Yamamura K. 1997. Efficiëntie van recombinatie door cre voorbijgaande uitdrukking in embryonale stamcellen: vergelijking van diverse promotors. J Biochem (Tokyo) 122: 977-82.
Brocard J, Feil R, Chambon P, Metzger D. 1998 a chimeric cre recombinase inducible by synthetic, but not by natural ligands of the glucocorticoid receptor. Nucleic Zuren Res 26: 4086-90.
Jax NOTES. 1999. NIH, Jackson Laboratory en DuPont Pharmaceuticals ondertekenen Cre-lox technologiegebruik overeenkomsten. JAX NOTES 476: 4.
Kellendonk C, Tronche F, Reichardt HM, Schutz G. 1999. Mutagenese van de glucocorticoïdereceptor bij muizen. J Steroid Biochem Mol Biol 69:253-9.
Nagy A. 2000. Cre recombinase: het universele reagens voor het aanpassen van het genoom. Genesis 26: 99-109.
Sadowski P. 1995. De FLP recombinase van het 2-µm plasmide van Saccharomyces cerevisiae. Prog Nucleïnezuur Res Mol Biol 51: 53-91.
Sauer B, Henderson N. 1988. Plaats-specifieke recombinatie van DNA in zoogdiercellen door de recombinase van Cre van bacteriofaag P1. Proc Natl Acad Sci U S A 85: 5166-70.
Sternberg N, Hamilton D. 1981. Bacteriofaag P1 plaats-specifieke recombinatie. I. recombinatie tussen loxp plaatsen. J Mol Biol 150: 467-86.
Utomo AR, Nikitin AY, Lee WH. 1999. Tijdelijke, ruimtelijke, en celtype-specifieke controle van CRE-bemiddelde recombinatie van DNA in transgene muizen. Nat Biotechnol 17: 1091-6.